庫爾勒香梨kfpSPL及其啟動子克隆分析
2018-08-30 11:39:04王玉凱牛建新
新疆農業科學 2018年6期
王玉凱,趙 歡,牛建新
(石河子大學農學院園藝系/特色果蔬栽培生理與種質資源利用兵團重點實驗室,新疆石河子 832003)
0 引 言
【研究意義】庫爾勒香梨是薔薇科植物、梨屬中的新疆梨種(Pyrussinkiangensisyu)。受遺傳特性的影響,庫爾勒香梨的果實萼片存在兩種情況,即脫落與宿存,萼片的宿存形成宿萼果,大量的宿萼果導致果實果形不規則,嚴重影響果實的口感、外觀與品質,致使庫爾勒香梨降低其商品價值和市場競爭力。從分子水平研究庫爾勒香梨kfpSPL基因與萼片脫落和宿存之間的關系,對提高庫爾勒香梨果實的品質及外觀有重要意義。【前人研究進展】近年來,關于庫爾勒香梨萼片的發育規律及其萼片脫落過程中相關內激素的變化規律已有相關報道[1,2]。前人研究表明,庫爾勒香梨的脫萼和宿萼差異表達的眾多片段中有兩條與蘋果中的轉錄因子有較高的同源性[3,4]。此外,孫曉霞等[5]運用RNA-Seq技術獲得庫爾勒香梨脫萼組和宿萼組代表樣品的轉錄本數據,篩選出脫萼組和宿萼組樣品的差異表達基因并且獲得了相應基因的功能注釋。在王博慧等[6]的研究中發現,通過克隆技術得到了kfpMYB及其啟動子,并對已獲得的上游序列進行了生物信息學分析。在植物開花調控過程中SPL轉錄因子具有重要意義,控制植物開花的光周期誘導途徑和赤霉素誘導途徑[7]。【本研究切入點】以往對于庫爾勒香梨脫萼和宿萼的研究主要集中在生理研究層面,如植物生長調節劑[8]、形態[1]、修剪[9]、光照[10]、授粉[11]等生理因素對庫爾勒香梨萼片脫落與宿存的研究。作為調控基因的重要組成部分,啟動子控制基因的表達時間和程度,它是一段特定的DNA序列,可與RNA聚合酶識別、結合及起始轉錄。研究庫爾勒香梨萼片脫落與宿存的分子機理。【擬解決的關鍵問題】克隆kfpSPL基因的全長基因組DNA序列及其啟動子,了解該基因如何應答激素和環境條件。
1 材料與方法
1.1 材 料
4月中旬在新疆庫爾勒市沙依東園藝場五分場,標記3株樹齡為10a的庫爾勒香梨樹作為試驗樹,于盛花期(全樹50%花朵開放),采集標記庫爾勒香梨樹的全花(花梗以上部分),三棵試驗樹各30朵花,去除花瓣并將剩余部分從萼片和子房交界處分割成兩部分,分別用錫箔紙包好,迅速投入液氮中保存,帶回實驗室置于-80℃超低溫冰箱中備用。
1.2 方 法
1.2.1 庫爾勒香梨kfpSPL的基因組DNA克隆
以天根公司新型植物基因組小型提取試劑盒說明書為依據進行庫爾勒香梨基因組DNA的提取。使用試驗已獲得的庫爾勒香梨kfpSPL基因的cDNA序列及梨基因組信息為模板設計引物。上游引物為SPL1-S為5‘-TTGCTGCCTCCTACCCTG-3’,下游引物為SPL1-A為5‘-ACTTGCCAAACTAAACATC-3’。以庫爾勒香梨基因組DNA序列為模板進行kfpSPL基因組DNA序列的擴增。其PCR反應體系為:10×LA PCR BufferⅡ(Mg2+Plus)2.5 μL,dNTP Mixture(2.5 mmol/L)4 μL,模板1 μL,上下游引物(20 μmol/L)1μL,LATaq酶(5 U/μL)0.25 μL,滅菌蒸餾水加至25 μL。PCR反應程序為:94℃預變性5 min;94℃變性30s;51℃退火30s;72℃延伸2 min;30個循環;72℃延伸10 min。
使用1%瓊脂糖凝膠電泳檢測PCR擴增產物的結果,使用膠回收試劑盒回收PCR產物。回收的PCR擴增產物與pEASY-T1載體連接,轉化至大腸桿菌DH5α感受態細胞中、使用麥康凱培養基進行篩選,通過菌液PCR篩選陽性克隆產物,將菌液PCR擴增的陽性克隆產物交由生工生物工程(上海)股份有限公司進行測序,測序結果使用DNAMAN進行分析。
1.2.2 kfpSPL啟動子片段的克隆
使用SPL基因cDNA全長與梨基因組數據庫進行比對,獲得SPL基因上游啟動子參考序列。利用參考序列設計引物,以庫爾勒香梨基因組DNA為模板經PCR擴增獲得庫爾勒香梨SPL基因上游約2 000 bp的序列。上游引物為SPL1-QDZ-S為5’-AAGGGCATGAACCATACGTACC-3’,下游引物為SPL1-QDZ-A為5’-TACCTGCCTTTAGCAGCGTT-3’。PCR反應體系同上,PCR反應程序為:94℃預變性5 min;94℃變性45s;53℃退火30s;72℃延伸2 min;30個循環;72℃延伸10 min。將PCR擴增產物片段連接到pEASY-T1克隆載體上,將已獲得的重組質粒交由生工生物工程(上海)股份有限公司進行測序,測序結果使用DNAMAN進行分析。
1.2.3 序列分析
將測序得到的kfpSPL基因組DNA序列和啟動子序列在梨全基因組序列數據庫(http://peargenome.njau.edu.cn)中進行比對,再在植物順式調控元件數據庫PLACE和PlantCare(http://bio_informatics.psb.ugent.be/webtools/plantcare/html/)中對啟動子序列進行順式作用元件預測分析。
2 結果與分析
2.1 kfpSPL基因的基因組DNA序列的克隆
使用引物SPL1-S和SPL1-A從庫爾勒香梨基因組DNA中擴增得到長度約為3 000 bp的片段,該片段經過克隆、菌液PCR驗證、挑選出陽性克隆,測序得到該片段長度為3 320 bp,與該基因的cDNA序列比較發現包含三段內含子,總長度為1 728 bp,第一段內含子位于基因組DNA的第407~949 nt處,第二段內含子位于第962~2 386 nt處,第三段內含子位于第2 530~2 673 nt處。
將獲得的kfpSPL基因的基因組DNA序列在梨全基因組數據庫中進行比對,發現該基因組DNA與Pyrus bretschneideri 數據庫中的一段序列的同源性為99%,且二者長度相同,E值為0,該序列位于scaffold291.0中301 708~305 026 nt的位點上,而另一段與Pyrus bretschneideri 數據庫中的一段序列的同源性為85%,且二者長度相同,E值為2.7,該序列位于scaffold291.0中300 845~300 878 nt的位點上。圖1
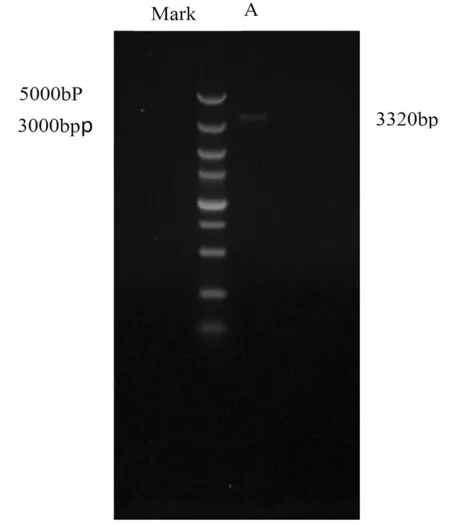
圖1 kfpPL基因啟動子的PCR擴增
Fig.1 PCR production of kfpSPL gDNA(A)
2.2 kfpSPL基因啟動子序列克隆
用kfpSPL基因的cDNA全長序列與梨基因組序列進行比對,獲得kfpSPL基因上游啟動子參考序列,根據比對后獲得的啟動子參考序列設計引物,通過PCR技術克隆獲得上游啟動子序列,經測序表明啟動子長為2 332 bp。將克隆得到的啟動子序列在梨全基因組序列數據庫中進行比對,發現克隆得到的上游啟動子序列的1~796 nt和1 042~2 332 nt分別與scaffold291.0中290 783~289 988 nt和289 990~288 701 nt的同源性為96%和97%,E值均為0。說明kfpSPL基因啟動子序列與碭山梨scaffold291.0啟動子序列存在一定的差異。其中位于kfpSPL基因的啟動子序列的第797~1 041 nt處與梨全基因組數據庫中未有比對上的相同序列,可能是因為庫爾勒香梨與碭山梨的品種間差異造成的。在kfpSPL基因的啟動子序列的第161~162 nt處發生插入突變,在1 345~1 350 nt處發生插入突變,在1 375~1 377 nt處發生插入突變,在2 129~2 137 nt處發生插入突變。并且在序列中多處有單堿基突變發生。圖2
2.3 kfpSPL基因啟動子序列
通過生物信息學分析kfpSPL基因的上游啟動子序列,其預測的轉錄因子結合位點及其功能。生物信息學分析結果表明,庫爾勒香梨kfpSPL基因的啟動子序列中的TATA-box和CAAT-box基本轉錄原件也存在于其他多數啟動子中,并分別位于kfpSPL基因啟動子序列上的多個位點,從預測中還發現一些順式調控元件,其中高轉錄水平的順式調控作用元件為5UTR Py-rich stretch,與光調控相關的順式作用元件ACE、Box 4、 Box II、G-box、CATT-motif和TCT-motif,與脅迫相關的順式作用調控元件有ARE、TC-rich repeats、WUN-motif,與脫落酸響應有關的順式作用調控元件是ABRE,與赤霉素響應有關的順式調控作用元件是GARE-motif、P-box,與水楊酸響應有關的順式調控作用元件是TCA-element,與分生組織表達相關的順式作用調控元件是CAT-box,與茉莉酸響應有關的順式作用調控元件是CGTCA-motif、TGACG-motif,與胚乳表達所需的順式作用調控元件是Skn-1_motif,MYB結合位點參與干旱誘導MYB的調控元件是MBS。這些調控元件的存在說明kfpSPL基因受多種外界信號的調控,例如激素、光、脅迫等。通過一系列的生物學過程來調控kfpSPL基因特定的轉錄過程。庫爾勒香梨kfpSPL基因的啟動子含有赤霉素、水楊酸、脫落酸等較為常見的順式作用元件,可以推斷赤霉素、水楊酸、脫落酸等激素可能會調控kfpSPL基因的表達。表1,圖3
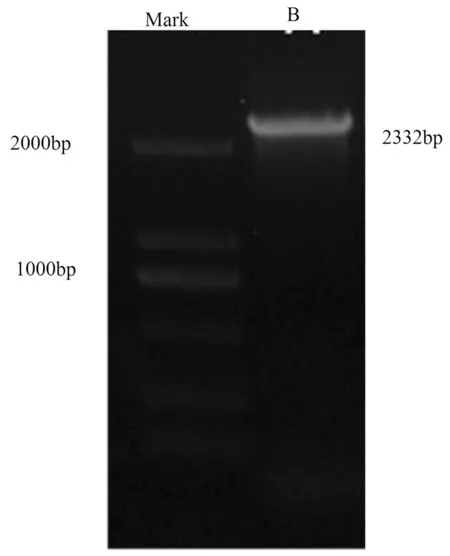
圖2 kfpSPL基因gDNA的PCR擴增
Fig.2 PCR production of kfpSPL promoter(B)
表1 應用 PLACE 和 Plant CARE 在線預測的啟動子區順式作用元件
Table 1 cis-acting regulatory elements analysis of promoter sequences by PLACE and plant CARE

調控序列名稱Regulatory sequence name位置(鏈)Location(chain)序列Sequence位點功能Fuction of site5’UTR Py-rich stretch-432TTTCTTCTCT高轉錄水平的順式作用元件cis-acting element conferring high transcription levelsAAGAA-motif-193GAAAGAABRE+1 155GCCACGTACA順式作用元件參與脫落酸的反應cis-acting element involved in the abscisic acid responsivenessACE+910ACGTGGA光響應中的順式作用元件cis-acting element involved in light responsivenessARE+108;-1 204;+1 020TGGTTT厭氧誘導必需的順式作用調控元件cis-acting regulatory element essential for the anaerobic inductionAT-rich element+536ATAGAAATCAA在豐富的DNA結合蛋白的結合位點(atbp-1)binding site of AT-rich DNA binding protein (ATBP-1)ATGCAAAT motif+365ATACAAAT順式調控元件相關聯的tgagtca模體cis-acting regulatory element associated to the TGAGTCA motifBox 4-916;-927ATTAAT參與光響應的保守DNA模塊的一部分part of a conserved DNA module involved in light responsiveness Box II+1 266TGGTAATAA光響應元件的一部分part of a light responsive elementCAAT-box+42等42個位點+42 ect,42sitesCAAT,CAATT,CCAAT啟動子和增強子區的共同順式作用元件common cis-acting element in promoter and enhancer regionsCAT-box+1 155GCCACT與分生組織表達相關的順式作用調控元件cis-acting regulatory element related to meristem expressionCATT-motif-1 352GCATTC光響應元件的一部分part of a light responsive elementCGTCA-motif-1 478CGTCA順式調控元件參與茉莉酸響應cis-acting regulatory element involved in the JAMe-responsivenessCTAG-motif+339ACTAGCAGAAAG-box+117;-678;-156;-678;-909CACATGG,TAAACGTG,CACGTC,CACGTT順式作用調節元件參與光響應性cis-acting regulatory element involved in light responsivenessGARE-motif+144AAACAGA赤霉素響應元件gibberellin-responsive elementMBS+28;+1 272;-1 064GAACTG,TAACTGMYB結合位點參與干旱誘導MYB binding site involved in drought-inducibilityP-box+1431GCCTTTTGAGT赤霉素響應元件gibberellin-responsive elementSkn-1_motif+662GTCAT胚乳表達所需的順式作用調控元件cis-acting regulatory element required for endosperm expressionTATA-box+15等28個位點+15 etc,28 sitesTATTTAAA,TATAAA,TATATAA,TATA,ATATAT,TATATAAATC,TTTTA核心啟動子元件約30轉錄起始core promoter element around -30 of transcription startTC-rich repeats-434;-606ATTTTCTTCA防御和應激反應中的順式作用元件cis-acting element involved in defense and stress responsivenessTCA-element-874GAGAAGAATA水楊酸反應中的順式作用元件cis-acting element involved in salicylic acid responsivenessTCCACCT-motif+528TCCACCTTCT-motif+1 097TCTTAC光響應元件的一部分part of a light responsive elementTGACG-motif+1 478TGACG順式調控元件參與茉莉酸響應cis-acting regulatory element involved in the JAMe-responsivenessUnnamed__1+911CGTGGUnnamed__3+911CGTGGUnnamed__4+217等12個位點+217 etc12sitesCTCCWUN-motif-899TCATTACGAA創傷響應元件wound-responsive element

圖3 kfpSPL啟動子序列TATA-BOX,CAAT-BOX(灰色標出),部分預測順式作用元件(黃色標出)
Fig.3 Sequence of kfpSPL gene promoter TATA-BOX and CAAT-BOX are black arrow.Part of the predicted cis-acting element is marked with a highlighted box

圖4 kfpSPL基因組DNA序列,翻譯起始密碼子(紅色粗體標出),內含子(紅色字體標出),外顯子(黑色字體)
Fig.4 Sequence of kfpSPL genomic DNA. The initial password of the translation is marked in bold red, The introns are highlighted in red, and the black font is exon
3 討 論
前人研究表明,庫爾勒香梨的脫萼果主要于初花期至盛花期形成,于末花期減弱[12],表明花期決定著果實果形的變化,這一研究與張大海等[13]研究結果一致。任瑩瑩等[14]認為,在初花期至花后兩周,尤其是盛花期,噴施NAA、PBO和乙烯利可影響庫爾勒香梨萼片宿存,其中噴施PBO可提高90%以上的脫萼率。由前人研究得知,使用PBO、ETH、NAA處理可增加庫爾勒香梨萼片的脫落[15],用GA處理可減少庫爾勒香梨萼片的脫落[2]。使用ABA、ETH和NAA三種激素處理使得庫爾勒香梨梨花kfpSPL基因的表達量上調,而用PBO和GA處理后kfpSPL基因表達量受處理時間和樹勢強弱的影響。kfpSPL基因很可能與庫爾勒香梨萼片脫落與宿存的激素調控有一定的關系。
4 結 論
kfpSPL基因啟動子序列中存在與赤霉素、脫落酸、水楊酸等激素相關的順式作用元件,不同生長調節劑處理脫萼果和宿萼果的花器官可以調控kfpSPL基因的表達水平。在kfpSPL基因的啟動子序列中可能含有對其在轉錄調控過程中起到重要作用的順式調控作用元件,如脫落酸響應式順式調控作用元件ABRE,赤霉素響應順式作用調控元件GARE-motif和P-box,水楊酸響應順式作用調控元件TCA-element,而這三種激素處理可能影響庫爾勒香梨萼片的脫落和宿存。
