高州油茶花芽分化與發育特征研究
2018-09-05 03:44:54奚如春黃容容
廣東農業科學 2018年7期
張 鵬,楊 穎,奚如春,2,黃容容,許 葉
(1.華南農業大學林學與風景園林學院,廣東 廣州 510642;2.廣東省森林植物種質創新與利用重點實驗室,廣東 廣州 510642)
油茶(Camellia oleiferaAbel.)為山茶科(Theaceae)山茶屬(Camellia)植物,是我國南方重要的木本油料樹種[1],與油棕(Elaeis guineensisJacq.)、椰子(Cocos nucifera L.)、油橄欖(Olea europaea L.)并稱為世界四大木本油料植物。高州油茶(Camellia gauchowensisChang ) ,又名大果油茶、華南油茶、越南油茶、陸川油茶,其樹體高大,果大葉茂,單位面積產量高,是山茶屬中一個較優良的物種[2-5]。
花是被子植物的重要特征之一,花芽分化是葉芽轉化為花芽發育成花器官雛形的重要階段[6-9]。花芽分化過程一般包括成花誘導、花芽發端、花芽形態建成或花芽發育階段[10]。對不同植物的花芽分化時期的劃分存在差異[11-13]。如早實核桃的花芽分化期可分為雌花花序分化期、花柄原基和雌花原基分化期、花被原基分化期、苞片原基分化期、花萼原基分化期、花瓣原基分化期、雌蕊原基分化期和胚珠分化期8個階段[14]。不同植物的花芽分化時期雖有差異,但總體來說,花芽分化的順序都是萼片、花瓣、雄蕊和雌蕊[15]。普通油茶在每年5~6月進入花芽分化期,持續到10月左右,因此整個花芽分化時間長。分化初期花芽分化快,但到了7~8月果實生長快速、內部物質積累多,花芽因營養不足導致大量脫落[16],9月中下旬大部分花芽的形態分化結束,此時花芽飽滿,10月下旬進入始花期,花芽分化結束。
花芽分化對植物開花、結果都有直接影響,目前油茶種植過程中普遍存在大量落花、落果問題,其花芽發育不良是導致油茶低產的重要原因之一[17],油茶花芽分化不同階段的外觀形態特征不同,花芽分化過程對果實的產量和質量有直接影響,通過控制花芽分化及果實的生長發育,可以達到優質豐產的目的[18]。目前對高州油茶的研究主要集中在常規栽培和引種上,對其花芽分化報道較少,在生產技術上存在爭議。本試驗以高州油茶為材料,觀察了花芽分化過程及其外部形態、含水率變化和養分含量變化,旨在闡明其花芽分化的生物學特征,對高州油茶生產具有很高的理論價值和現實意義,為科學合理的栽培技術奠定基礎。
1 材料與方法
1.1 試驗地概況
試驗地位于廣東省揭陽市揭東區新亨鎮新西河水庫庫區揭陽市林科所油茶試驗地。該地屬亞熱帶季風氣候,日照充足、雨量充沛。試驗林總面積30 hm2,為20世紀70年代初采用高州油茶實生苗栽植的人工林,林地總體坡向西北、坡度30°~40°,土壤類型為山地黃紅壤。目前該林分管理條件好,總體生長良好。
1.2 樣地設置及樣品采集
在試驗區內進行全林踏查后,為使采樣樹具有隨機性和代表性,分別在上、中、下坡選擇具有代表性的地段內設置3個樣地,每個樣地面積為20 m×20 m(有坡度時進行改平);在每個樣地內選擇生長旺盛、無病蟲害的采樣樹10株,共30株。
于2016年5~10月進行野外樣品采集及花芽分化期觀測。分別于每月的上、中、下旬進行采樣,每隔10 d采樣1次,每次隨機采集60個大小相似、發育進程一致的花芽,其中30個立即用FAA(福爾馬林5 mL+冰醋酸5 mL+70%酒精90 mL)固定保存,30個立即帶回實驗室處理。
1.3 試驗方法
1.3.1 花芽外部形態觀察測定 用游標卡尺測量花芽的長度和寬度,精確至0.01 mm,并觀察記錄其外部形態特征。
1.3.2 含水率測定 稱量花芽的鮮重后,置于65℃ 烘箱恒溫烘24 h以上(至恒重),冷卻后及時稱量干重,計算其含水率。
1.3.3 花芽養分含量測定 將烘干后的樣品貼好標簽干燥保存,測定花芽的N、P、K、Ca、Mg、Zn、B等養分含量。全氮采用硫酸-雙氧水消煮-蒸餾滴定法;全磷采用硫酸-雙氧水消煮-釩鉬黃比色法;全鉀采用硫酸-雙氧水消煮-火焰原子吸收分光光度法;鈣、鎂、鋅采用干灰化-稀鹽酸溶解-火焰原子吸收分光光度法;硼采用干灰化-稀鹽酸溶解-姜黃素比色法。
1.3.4 常規石蠟切片制作與觀察 參照李和平等[19]的常規石蠟切片制作方法。具體試驗步驟如下:固定,脫水,透明,浸蠟和包埋,切片,展片、粘片及烤片,脫蠟,復水,番紅—固綠染色,脫水,透明,封片。使用OLYMPUS BX-51型顯微鏡觀察花芽縱切面的內部解剖結構,并且拍攝照片,記錄花芽分化過程中內部結構的變化特征。
試驗數據采用Microsoft Excel 2007進行分析。
2 結果與分析
2.1 花芽的解剖結構及外觀形態特征
對試驗地的高州油茶進行觀測結果表明,其樹體每年3月中旬開始萌發春梢,5月中旬少量林緣樹體的花芽開始分化,5月下旬大部分樹體的花芽開始分化,光照不足的樹體進入花芽分化的時間稍晚,個體間存在差異。由于每年的氣溫、濕度等差異,花芽分化也略有差異。根據花芽分化特征,本文將高州油茶花芽分化分為6個時期,即前分化期、萼片形成期、花瓣形成期、雌雄蕊形成期、子房與花藥形成期、雌雄蕊成熟期,各個時期的解剖結構及外觀形態見圖1(封二)。
2.1.1 前分化期 5月中旬進入前分化期(圖1a2),此時其生長點開始發生變化,分裂較快,初期生長點較尖,后期逐漸變圓,生長錐增大呈半圓形或圓錐形,前分化期為5月中旬至6月上旬。花芽外觀形態(圖1a1,封二):此時期花芽很小,長2.52~4.21 mm,寬2.06~2.88 mm,外觀形態與葉芽無明顯區別,很難區分,后期花芽逐漸增寬。
2.1.2 萼片形成期 6月上旬進入萼片分化期(圖1b2,封二),約持續15~20 d,分化前期生長點周圍出現花萼原基小突起,逐漸伸長并向內彎曲,后期覆蓋生長點,萼片形成期為6月上旬至6月下旬。花芽外觀形態(圖1b1,封二):長 3.00~5.03 mm,寬 2.21~3.46 mm,形狀逐漸變寬,外觀較扁,左右可見苞片2~3片。
2.1.3 花瓣形成期 6月下旬至7月中旬進入花瓣形成期(圖1c2,封二)。在萼片形成期的末期,生長點開始出現花瓣原基,花芽進入花瓣分化期。花瓣原基逐漸向上生長,縱向生長比橫向生長稍快,頂端較圓,花瓣原基最后分化成花瓣。花芽外觀形態(圖1c1,封二):此時花芽膨大速度較快,長3.70~7.48 mm,寬2.52~3.87 mm ,形態較為圓胖,可見3~4片苞片,與葉芽區分明顯。
2.1.4 雌雄蕊形成期 7月中旬,在花瓣形成期后期,生長點變得更寬, 在生長點上出現雌雄蕊原基小突起,此時為雌雄蕊形成期(圖1d2),由7月中旬至8月中旬,大約20~30 d。中間較大的突起為雌蕊原基,后期雄蕊原基在雌蕊原基附近生長。花芽外觀形態(圖1d1,封二):花芽生長快速,長度增長較快,長7.77~16.21 mm,寬 4.43~6.37 mm,新增苞片被有少量絨毛,可見4~5片苞片。
2.1.5 子房與花藥形成期 8月中旬至9月中旬分化,在雌雄蕊形成后期,雌蕊原基下部膨大,形成子房,雌蕊原基頂部逐漸靠攏形成柱頭和子房室,子房室內著生有多個胚珠,同時雄蕊原基分化形成花藥(圖1e2,封二)。花芽外觀形態(圖1e1,封二):花芽持續增大,長11.71~16.47 mm,寬 5.76~6.98 mm,此時寬度增長較快,外觀圓胖,苞片上被有大量絨毛。
2.1.6 雌雄蕊成熟期 9月下旬到10月中旬分化,20~30 d,最晚的樹體至10月下旬左右。此時期花柱繼續伸長,子房持續膨大,內含3~5室,每室兩個至多個胚珠,此時花藥已全部形成,花藥囊內已有花粉母細胞開始分化。10月中下旬子房內的胚珠基本發育成型,有內含物,花藥內已有大量花粉(圖1f2,封二)。花芽外觀形態(圖1f1,封二):外形由長度增長轉變為寬度增長,長13.73~18.75 mm,寬6.53~9.00 mm,形態圓胖,顏色逐漸由嫩綠色變為黃綠色,內部的花藥由無色逐漸變成淺黃色。
2.2 花芽長寬比變化特征
高州油茶花芽長寬比(長/寬)在花芽分化時期內的變化如圖2所示。整個花芽分化期內,花芽不斷增大,花芽平均長度由5月中旬的3.55 mm增長到10月下旬的17.44 mm,平均寬度由2.63 mm增大到7.56 mm。從圖2可以看出,花芽長寬比總體表現為先緩慢增長,經歷短暫的快速增長期后又保持平緩增長趨勢。
5月15日至6月5日,花芽處于前分化期,花芽體積逐漸變大,花芽長寬比略有增大,由1.35增大到1.48。6月5日后,花芽分化處于前分化期的末期,此時花芽分生組織分裂加快,花芽橫向生長快于縱向生長,增寬明顯,長寬比有所下降。6月15日之后,花芽分化處于萼片形成期的后期,此時縱向生長加速,長寬比開始回升。6月25日開始,花芽進入花瓣形成期,此時橫向增長快于縱向生長,長寬比從1.65下降到1.53,外觀表現圓胖。在7月15日花芽進入雌雄蕊形成期,雌雄蕊原基縱向生長加快,花芽長寬比急劇增加,由1.53迅速增長到2.13,后趨于平緩。8月15日花芽開始進入子房與花芽形成期,前期縱向生長依舊保持較快速度,長寬比有一次上升,8月25日時達到整個分化時期的最大值2.33,后由于子房開始膨大,長寬比下降,9月5日為2.23。進入雌雄蕊成熟期后,伸長和增寬同時進行,長寬比無明顯變化。整個分化過程花芽的長寬比經歷增大、減小、增大再減小,最后趨于平緩的過程,這與花芽分化過程中各個組織的生長密切相關。

圖1 花芽分化外觀形態及其對應內部解剖結構
2.3 花芽含水率變化
高州油茶在花芽分化時期內花芽含水率隨時間的變化如圖3所示。整個分化時期花芽含水率在41.51%~55.32%之間,在分化期內經歷下降、上升、下降的反復過程,總體表現為從花芽分化開始到結束時花芽含水率有所降低。
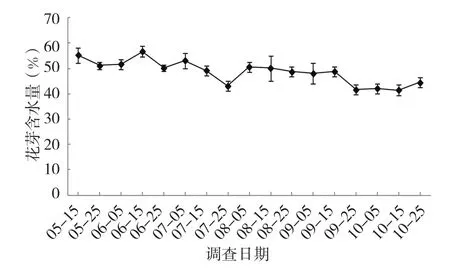
圖3 花芽含水率的動態變化
5月15~25日,花芽含水率由55.32%下降到51.10%,此時花芽剛開始進入前分化期,組織開始加快生長,營養物質增多,水分含量略有降低。6月上旬,進入萼片形成期,花芽含水率經歷先上升后下降的過程。隨后進入花瓣形成期,含水率先上升2.98%,花瓣形成期后期花芽含水率急劇下降,從7月5~25日持續下降,由53.17%下降到43.07%,下降了10.10%,此時花芽膨大速度較快,營養物質積累快速,消耗一定的水分。雌雄蕊形成后期,含水率略有提升,8月5日的含水率上升為50.64%。8月中旬至9月中旬,處于子房與花藥形成期,含水率沒有明顯變化。之后進入雌雄蕊形成期,含水率有所下降,降幅為7.21%,隨后雌雄蕊分化基本完成,含水率較為穩定,波動不大。
2.4 花芽養分含量變化
高州油茶花芽在花芽分化時期全N、全P、全K、全Ca、全Mg、全Zn、全B的含量變化如圖4和圖5所示。

圖4 花芽中N、P、K、Ca、Mg含量的動態變化
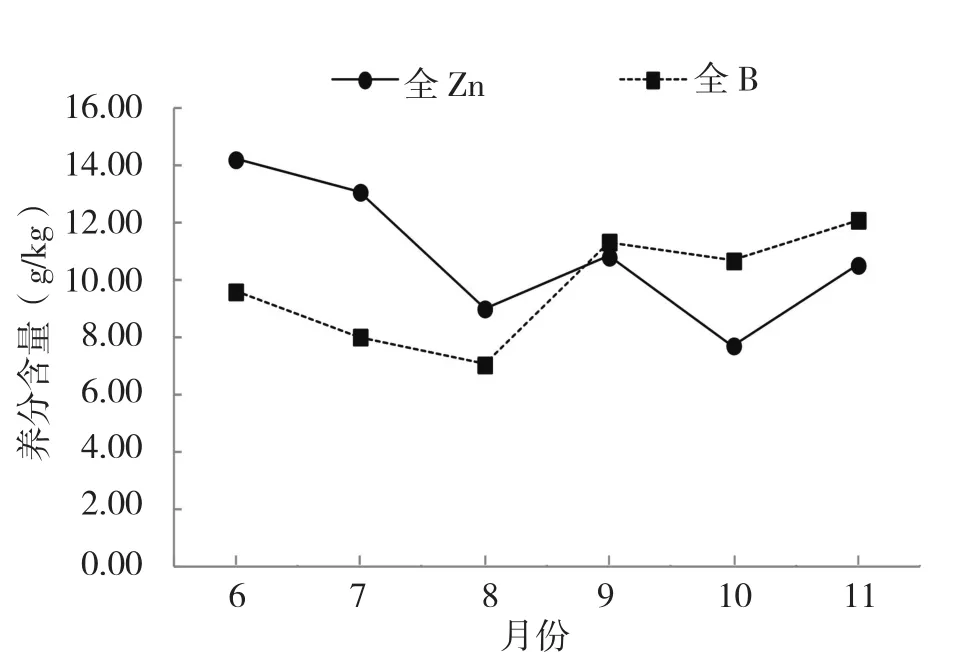
圖5 花芽中Zn、B含量的動態變化
全N含量在6~8月變化較為平緩,總體上略有下降,這段時期花芽中的組織生長旺盛,可能消耗了花芽中原有的蛋白質轉變為其他物質,8月是整個時期的最低值6.37 g/kg,隨后全N含量持續上升,到11月達到最大值9.93 g/kg。全K含量在6~7月略有下降,但降幅不大,只下降0.39 g/kg,之后在7~11月全K含量逐漸上升,到11月達到最大值6.63 g/kg,碳水化合物在植物花芽形成過程中起到重要作用,而K對碳水化合物的合成和轉移有重要作用,全K含量持續上升,促進花芽分化。全Ca含量在6~11月總體表現出下降的趨勢,特別是6~8月花芽分化前期,全Ca含量急劇下降,由8.63 g/kg下降到3.62 g/kg,花芽分化時消耗大量的Ca,最后維持一個適度的水平。全P和全Mg含量在花芽中的含量較低,在整個花芽分化時期的變動不大,均在1.0 g/kg上下波動。全Zn含量在整個時期的波動很大,在6~8月急劇下降,由14.23 mg/kg下降到9.01 mg/kg,Zn在蛋白質合成中起到不可或缺的作用,在花芽快速分化階段Zn的消耗較大。隨后,全Zn的含量略有上升,但9~10月雌雄蕊形成期其含量又從10.82 mg/kg下降到7.72 mg/kg,10~11月,全Zn的含量回升至10.54 mg/kg。全B含量在6~8月略有下降,但在8~11月持續上升,尤其在8~9月上升速度加快,從7.06 mg/kg上升到11.35 mg/kg。全B含量在分化前期低于全Zn含量,但在9月份之后,全B含量高于全Zn含量。花芽分化結束時,花芽中養分含量的高低依次為:全N>全K>全Ca>全 P>全 Mg>全 B>全 Zn。
3 結論與討論
高州油茶的花芽分化時間長,于每年5月中旬至10月中旬進行花芽分化,持續時間100~120 d,比袁德義等[20]觀察到的湘林 1號的143 d略短。花芽分化可分為6個時期:前分化期、萼片形成期、花瓣形成期、雌雄蕊形成期、子房與花藥形成期、雌雄蕊成熟期,與莊端林[21]對湘林系列油茶、劉曲等[22]對浙江紅花油茶、朱雯等[23]對廣寧紅花油茶花芽分化時期的劃分結果一致。花芽分化的每個時期持續的時間不同,其外觀形態和內部特征變化明顯。在觀察中發現,各單株花芽分化的時間有差異,不是所有花芽都集中在短期內進行花芽分化,而是分期、分批進行,同一時期在不同單株上采集的花芽可能發育進程相差很大,總體表現為光照充足的樹體進入花芽分化的時間較早。說明光照充足有利于花芽提早分化,在生產中可適當調整林分密度與樹體管理進行花期調控。
長寬比的變化與花芽各個組織的生長密切相關。花芽分化時,長寬比總體變化特征為:先緩慢增長,經歷短暫的快速增長期后又趨于平緩。花芽含水率總體表現為從花芽分化開始到結束時花芽含水率有所降低。含水率的下降可能是因為花芽分化時內部的營養物質增多,水分在總鮮重的比例有所降低。高州油茶在進行花芽分化時,花芽的養分元素含量有所變化。全N含量總體表現為先下降后上升,N是蛋白質的重要組成成分,全N含量的變化與花芽分化時蛋白質的消耗和合成密切相關。全K含量在整個時期逐漸上升,K對碳水化合物的合成和轉移有重要作用,全K含量持續上升,促進了花芽分化。全Ca含量總體表現為下降趨勢,花芽分化時消耗大量的鈣,最后維持一個適度的水平。鈣在植物花芽分化中起重要作用,促進植物的花芽分化,彭抒昂等[24]對梨成花過程中鈣的動態變化研究中也發現Ca含量在花芽發端時出現高峰值,隨著花芽分化的進行逐漸被消耗。全P和全Mg含量在花芽中的含量較低,在整個花芽分化時期的變動不大。全Zn含量在花芽分化中的波動很大,總體表現為先下降、后上升、再下降、再上升的過程,Zn對植物的生長發育有著良好的促進作用,在整個花芽分化過程中與其內部組織的生長發育密切相關。花芽中全B的含量總體表現為先下降后上升。B能與細胞壁成分組成復合體,是細胞壁半纖維素的組成成分,同時B集中存在于植物的根尖、莖尖和花器官中,所以與植物的花芽分化和花粉萌發密切相關[25]。花芽分化發育過程對果實的產量和質量有直接影響,通過養分控制,調控花芽分化及發育,實現高州油茶優質 豐產。
