枳NLP轉錄因子響應干旱脅迫并與NRE順式作用元件互作
2018-09-11 06:19:18曹雄軍盧曉鵬熊江李靜謝深喜
中國農業科學 2018年17期
曹雄軍,盧曉鵬,熊江,李靜,謝深喜
(1湖南農業大學園藝園林學院/國家柑橘改良中心長沙分中心,長沙 410128;2廣西壯族自治區農業科學院葡萄與葡萄酒研究所,南寧 530007)
0 引言
【研究意義】我國柑橘產業主要分布在南方丘陵山地,容易受到土壤干旱脅迫的影響。即使我國南方濕潤地區也經常會在 7—9月份發生區域性和季節性的干旱[1],此時正值柑橘果實膨大及品質產量形成的關鍵時期,如橘園不能有效地通過灌溉來解決干旱問題,則會給柑橘的安全生產帶來較大壓力。干旱脅迫影響氮素營養的吸收、運轉和代謝[2]。氮素是作物生長發育所必需的大量元素之一,是植物體內許多重要有機化合物的組分,參與和調節植物在干旱逆境下的適應、傷害、修復和補償等生理生化過程[3-4]。筆者所在研究團隊前期研究表明,隨著干旱脅迫的加劇,柑橘葉片葉綠素含量下降,柑橘體內吸收的氮素含量減少[5]。目前,隨著全球氣候變暖和生態環境的惡化,干旱發生的頻率和強度均有所加劇[6-7],干旱脅迫給柑橘安全生產帶來的壓力越來越突出。柑橘的水分生理和營養吸收研究密不可分,在干旱條件下,研究柑橘對干旱脅迫的響應,以及氮素代謝機制等,對實際生產中調控柑橘生長發育、品質與產量具有重要意義。【前人研究進展】柑橘屬于典型的亞熱帶樹種,喜溫暖、濕潤的生態環境,對水分有較高的需求[8]。水分的缺失會使柑橘的形態發生變化,如植株生長緩慢或停止、葉片和嫩莖萎蔫、葉片枯死和脫落;使葉片氣孔開度減小甚至關閉[9-11],與光合作用有關的細胞器受損,導致葉片光合速率下降[12-15];也影響根系對礦質元素(N、P、K 等)吸收[5,16-17]。因此,干旱脅迫對柑橘的生長發育、果實品質與產量具有極為不利的影響[18]。研究發現,豆科植物根瘤形成依賴于起始結瘤基因(nodule inception,NIN)的存在,而在非豆科植物中,也有類似基因存在,參與硝酸鹽的同化,命名為NLP(NIN-like protein)基因[19]。研究表明,擬南芥(Arabidopsis thaliana)NLP轉錄因子不僅在調控氮素吸收與同化中扮演著非常重要的角色[20-23],而且與擬南芥的抗旱性有關[24]。NIN-like家族蛋白具有感知硝酸鹽信號的能力,能與硝酸鹽響應順式作用元件(nitrate-responsive cis-element,NRE)綁定,激活依賴NRE啟動的基因表達[20-21]。另外研究人員發現,敲除了AtNLP7的擬南芥相比正常野生型表現出了較強的抗旱能力[24]。【本研究切入點】到目前為止,關于柑橘 NLP轉錄因子參與氮素吸收與同化還未有深入研究,因此,本文針對枳(Poncirus trifoliata(L.)Raf.)氮素含量受干旱的影響,探討枳NLP轉錄因子對干旱脅迫的響應,以及其與NRE的互作關系。【擬解決的關鍵問題】分析干旱條件下枳葉片和側根中氮素含量的變化以及枳亞硝酸還原酶基因(PtNiR)和NLP轉錄因子的表達,克隆PtNiR啟動子序列并進行順式作用元件分析,運用酵母單雜交技術分析驗證枳NLP轉錄因子與NiR啟動子上硝酸鹽響應順式作用元件的互作關系。
1 材料與方法
試驗于 2015—2017 年在湖南農業大學國家柑橘改良中心長沙分中心進行。
1.1 試驗材料
試驗材料為枳播種實生苗,種子來源于湖南省洞口縣洞口鎮大田村同一棵母本株‘大田1號’,遺傳基礎一致。枳實生苗栽培于溫室大棚中的塑料盆中(直徑25 cm,高度23 cm),其中試驗植株108株,樹齡為1年生并生長一致,栽培基質由草炭、鋸木屑及細河沙按照1∶2∶1的比例混勻而成。
1.2 試驗方法
1.2.1 枳干旱處理及采樣 以植株統一澆透水為處理第1天,此后每天在上午9:00—10:00用土壤水分測量儀(TDR300,USA)監測栽培基質的持水量。干旱組的基質不再補充水分直到觀察到植株葉片萎蔫,而對照組的基質相對持水量下降至55%—65%時,每2 d補充適量水分使對照基質保持在該范圍。處理期間采集干旱組和對照組試驗分析樣本3次,即統一澆水后第7天基質相對持水量在55%—65%范圍時采樣一次;第27天干旱組基質相對持水量在18%—22%時采樣一次;第37天干旱組觀察到植株葉片萎蔫、基質相對持水量在12%—16%時采樣一次,每次采樣干旱組和對照組分別采集3組重復,每組6株,其中3株采集新梢頂部3—5片葉,以及根部幼嫩側根,用于RNA提取和熒光定量PCR分析;另外3株采集全部葉片和側根用于氮素含量的測定。
1.2.2 氮素含量的測定 將待測枳葉片、側根樣品取好后,立即用水洗凈,在烘箱中設置15 min 120℃,然后于65℃條件下烘干至恒重,研磨成粉后采用凱氏定氮儀測定全氮含量。
1.2.3 枳DNA提取 用液氮研磨葉片,參照CTAB法提取枳 DNA。使用超微量分光光度計(Implen P330,德國)檢測 DNA濃度與純度,同時用 1.0%的瓊脂糖電泳檢測DNA的完整性,檢測合格后,貯于-20℃備用。
1.2.4 RNA提取及 cDNA制備 使用 Trizol試劑盒(Invitrogen,USA)參照說明書的步驟提取枳葉片和側根的總RNA,之后在超微量分光光度計(Implen P330,德國)上檢測RNA濃度與純度,同時用1.0%的瓊脂糖電泳檢測總RNA的完整性,檢測合格后,使用反轉錄試劑盒(PrimeScriptTMRT reagent Kit with gDNA Eraser(Perfect Real Time),TaKaRa)制備實時熒光定量分析cDNA模板,完成后于-20℃保存備用。
1.2.5 引物設計 參照甜橙基因組數據庫序列,使用Oligo 7.0軟件設計枳NLP基因全長的PCR擴增引物[23],設計PtNiR的啟動子擴增引物;使用Beacon Designer 8設計實時熒光定量PCR引物(表1)。
1.2.6 熒光定量PCR分析 儀器采用BIO-RAD公司的CFX96 real-time PCR儀。擴增條件設置為兩步法:95℃預變性30 s后,95℃變性5 s,58℃ 15 s,40個循環,每個樣品 3次重復。選擇 YAN等[25]報道的ActinB作為內參基因,分別以對照樣品為參照分析基因的差異表達。根據LIVAK等[26]報道的2-ΔΔCT法分析枳葉與側根的qPCR數據,并使用SPSS軟件對基因相對樣品的表達數據進行差異顯著性分析。
1.2.7 基因克隆與酵母單雜交互作載體構建 使用高保真 DNA聚合酶(PrimerSTAR Max DNA Polymerase,TaKaRa)PCR擴增枳 NLP轉錄因子及PtNiR啟動子,擴增的目的產物經1%的瓊脂糖凝膠電泳后,把預期片段大小的目的條帶切膠、回收純化后,使用必克隆試劑盒(Ligation-Free Cloning Kit,abm),按照說明書進行連接轉化,篩選培養,然后提取載體質粒進行測序驗證。枳4×NRE序列由鉑尚生物技術(上海)有限公司合成并連接至pHIS2載體。
1.2.8 酵母單雜交分析 按照酵母單雜交步驟將構建的pHIS2-4×NRE和pGADT7-Rec-NLP載體質粒共同轉化到酵母菌株 Y187中,在氨基酸缺失培養基 SD/-Trp/-Leu(對照)以及 SD/-His/-Trp/-Leu/+120 mmol·L-13-氨基三唑(3-aminotriazole,3-AT)上30℃恒溫培養3—5 d,觀察酵母的生長情況,以共轉化pHIS2-4×NRE和pGADT7-Rec空載體的Y187酵母為陰性對照。
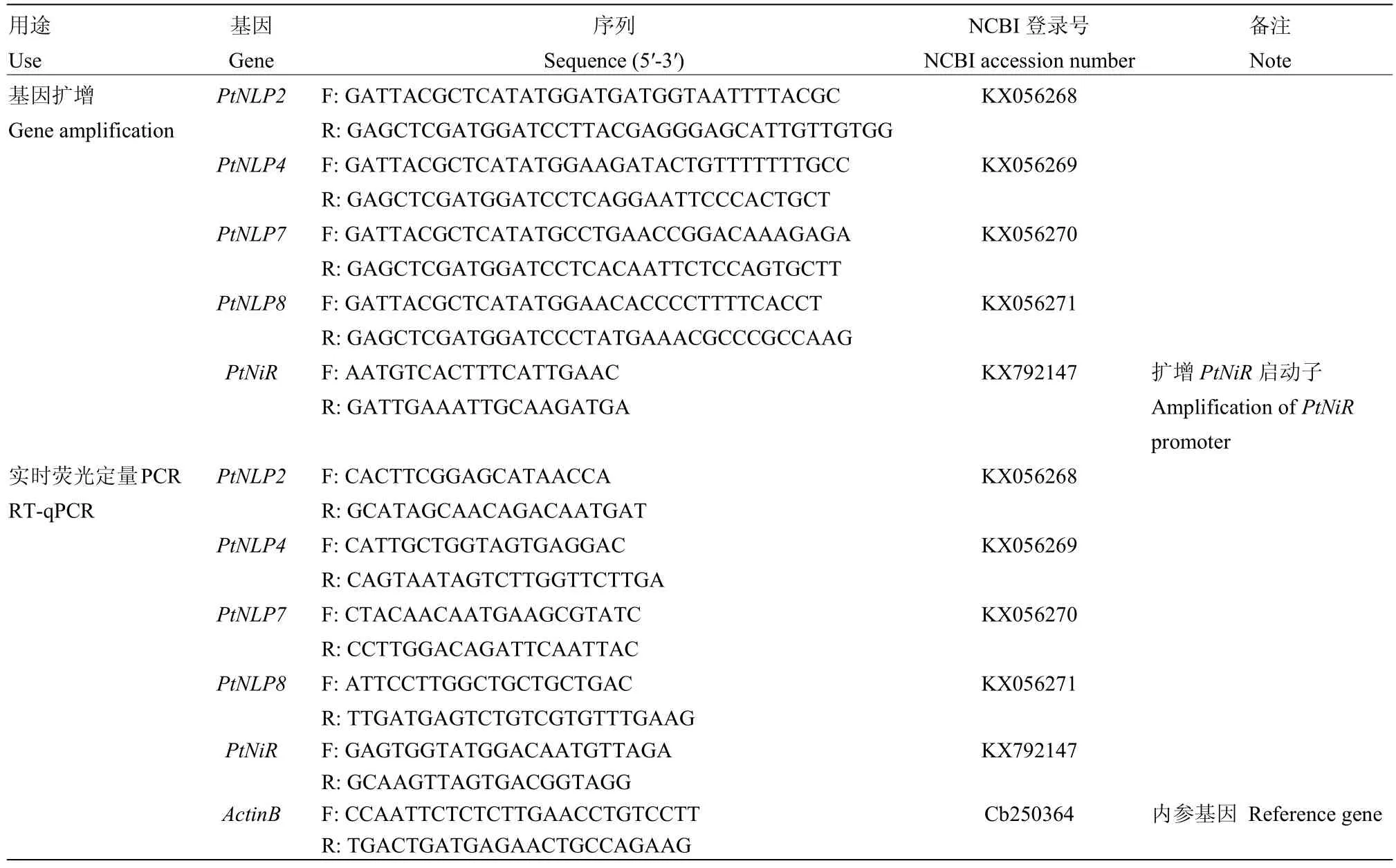
表1 本試驗設計的引物Table 1 Primers designed in this study
2 結果
2.1 干旱條件下枳氮素含量的變化
在干旱組中,葉片全氮含量表現出先升高再下降的趨勢,而側根中全氮含量顯著下降;在對照組中,枳葉片和側根全氮含量均顯著上升(圖 1)。以對照為參比,全氮含量在枳葉片和側根中均隨著干旱時間的延長而相對減少。
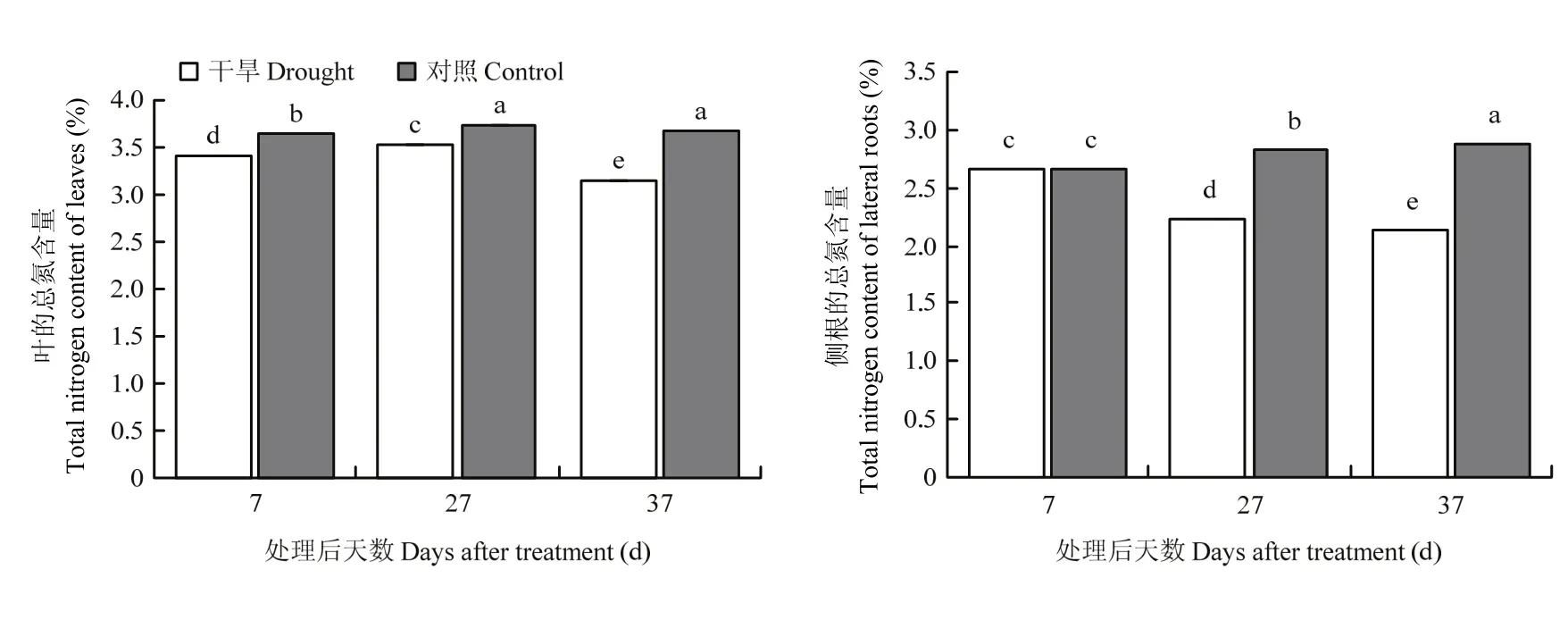
圖1 干旱處理后枳葉和側根中的全氮含量Fig. 1 The total nitrogen content in leaves and lateral roots of P. trifoliata after drought treatment
2.2 干旱條件下PtNiR的表達
分析了干旱條件下亞硝酸還原酶PtNiR(NCBI登錄號 KX792147)在葉片和側根的表達情況,結果表明,當第27天基質相對持水量在18%—22%時,葉PtNiR的表達水平上調超過處理第7天,但不顯著,而側根PtNiR的表達水平有所下調且顯著低于處理第7天;當第37天觀察到植株葉片萎蔫,基質相對持水量在12%—16%范圍時,葉和側根PtNiR的表達水平均顯著下調至更低的水平(圖2)。

圖2 枳葉和側根中PtNiR的相對表達量Fig. 2 The relative expression of PtNiR in leaves and lateral roots of P. trifoliata
2.3 干旱脅迫下枳NLP基因的表達
分析了本試驗條件下PtNLP2(NCBI登錄號:KX056268)、PtNLP4(NCBI登錄號:KX056269)、PtNLP7(NCBI登錄號:KX056270)及PtNLP8(NCBI登錄號:KX056271)在葉片和側根的表達量(圖3、圖 4)。結果發現,在葉片中,PtNLP2、PtNLP4及PtNLP7的表達趨勢相似,干旱第27天(基質相對持水量在18%—22%時)的表達量相對干旱第7天(基質相對持水量在55%—65%時)顯著上調,到干旱第37天(植株葉片萎蔫,基質相對持水量在12%—16%時),它們的表達量有所下調,其中,PtNLP4、PtNLP7下調至干旱第7天的表達量水平以下,但PtNLP2仍顯著高于干旱第7天。PtNLP8的表達量在干旱第27天有所下調,低于第7天的表達量但無顯著差異。
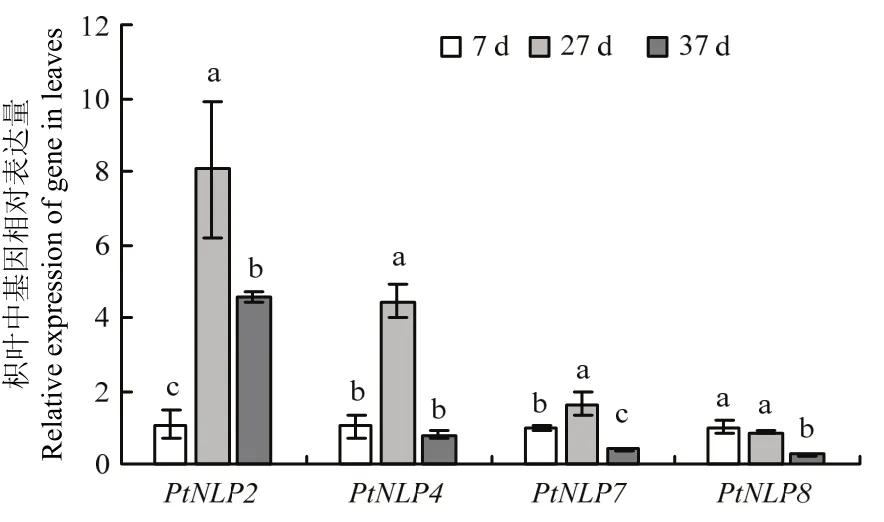
圖3 枳葉中NLP基因的相對表達量Fig. 3 The relative expression of NLP genes in leaves of P.trifoliata
而在枳側根中,同樣,PtNLP2、PtNLP4及PtNLP7在干旱第27天(基質相對持水量在18%—22%時)的表達量相對干旱第 7天(基質相對持水量在 55%—65%時)上調,其中PtNLP2、PtNLP7顯著上調;到干旱第37天(植株葉片萎蔫,基質相對持水量在12%—16%時),它們的表達量有所變化,其中,PtNLP2表達下調但仍顯著高于干旱第7天,PtNLP4表達下調至干旱第7天的表達水平以下,PtNLP7表達與干旱第7天無顯著性差異。PtNLP8的表達量在干旱第27天、第37天相比第7天均有所下調但無顯著差異。
從基因表達模式的角度分析,PtNLP2、PtNLP4及PtNLP7之間存在相似性,但與PtNLP8存在差異。NLP與PtNiR表達模式上具有一定的相關性,表明枳NLP轉錄因子與PtNiR表達可能存在關聯,因此,克隆分析了PtNiR的啟動子序列,并發現一段疑似NRE序列。

圖4 枳側根中NLP基因的相對表達量Fig. 4 The relative expression of NLP genes in lateral roots of P. trifoliata
2.4 PtNiR啟動子的克隆及序列分析
通過克隆測序獲得了包括 1 602 bp啟動子的PtNiR序列(NCBI登錄號KX792147)。通過檢索比對分析,在PtNiR啟動子區域-196至-154的位置發現了疑似 NRE,該 NRE由一個高保守 motif(TG(A/G)C(C/T)CTT)以及另一個低保守motif組成(圖 5-A、5-B),枳 NRE的第一個 motif序列為“TGACCCTT”,第二個 motif序列為“AAGAGTCC”,與菜豆(Phaseolus vulgaris)、菠菜(Spinacia oleracea)一致,與擬南芥、白樺(Betula pendula)、玉米(Zea mays)、高粱(Sorghum bicolor)、水稻(Oryza sativa)類似(圖5-A)。
2.5 枳NLP轉錄因子與NRE互作分析

圖5 硝酸鹽響應順式作用元件(NRE)Fig. 5 Nitrate-responsive cis-element (NRE)
為分析和驗證枳NLP轉錄因子與NRE的互作,構建了含有4×NRE-HIS3報告基因的pHIS2載體,并分別構建了融合PtNLP2、PtNLP4、PtNLP7及PtNLP8的表達載體pGADT7-Rec。分別將pHIS2-4×NRE-HIS3和 pGADT7-Rec-NLP2、pHIS2-4×NREHIS3和pGADT7-Rec-NLP4、pHIS2-4×NRE-HIS3和pGADT7-Rec-NLP7、pHIS2-4×NRE-HIS3和pGADT7-Rec-NLP8、pHIS2-4×NRE-HIS3和 pGADT7-Rec共同轉化至酵母菌株Y187中,生長在對照(/-Trp/-Leu)及篩選(/-His/-Trp/-Leu/+120 mmol·L-13-AT)培養基上。結果如圖6所示,轉化pHIS2-4×NRE-HIS3載體和不含NLP的pGADT7-Rec空載體的Y187酵母在對照培養基(-Trp/-Leu)能正常生長,而在含有120 mmol·L-13-AT的三缺培養基(-His/-Trp/-Leu)上不生長,說明酵母單雜交系統工作正常;轉化pHIS2-4×NRE-HIS3載體和pGADT7-Rec-NLP載體的Y187酵母在對照培養基(-Trp/-Leu)和含有120 mmol·L-13-AT的三缺培養基(-His/-Trp/-Leu)上均能正常生長。該結果表明在Y187酵母中,PtNLP2、PtNLP4、PtNLP7及PtNLP8均能與NRE綁定互作,暗示枳NLP轉錄因子能夠通過綁定NRE結合位點調控PtNiR表達。

圖6 酵母單雜交分析枳NLP轉錄因子與NRE的互作關系Fig. 6 Analysis of interaction between NLP transcription factor and NRE in P. trifoliata by Y1H assay
3 討論
研究表明,受干旱脅迫的影響,山下紅溫州蜜柑和紐荷爾臍橙的氮素吸收量顯著低于正常澆灌處理,且隨著土壤干旱程度的加深而減少[5]。本試驗的全氮含量結果表明,從處理后第7天到葉片開始萎蔫的第37天,無論是葉片,還是與氮素吸收轉運直接相關的側根,其本身的全氮含量均顯著減少,且相比對照,其相對含量也顯著減少。因此可見,干旱脅迫顯著影響柑橘的氮素營養,這可能與干旱脅迫能誘導或抑制氮代謝相關基因的表達有關。
硝酸鹽是高等植物中的主要氮源,也起到誘導生長和基因表達變化的信號分子的作用。硝酸鹽的同化作用是一個受到高度調控的過程,其受到氮素代謝如亞硝酸還原酶(NiR)等相關基因表達的影響[19,27-28]。本研究分析結果表明,枳側根中的亞硝酸還原酶PtNiR表達受到干旱脅迫的抑制,且隨著干旱時間的延長其表達量顯著下調。然而,PtNiR表達變化可能是受到干旱脅迫的直接影響,也可能是受到其他轉錄因子的調控。氮素代謝途徑中存在一些在硝酸鹽吸收以及同化的過程中起重要調控作用的轉錄因子[20-22,29-31]。研究發現,擬南芥中 NLP轉錄因子的活性可以受硝酸鹽信號的調節,其蛋白能與硝酸鹽響應順式作用元件(NRE)綁定結合,從而激活相關基因的表達[20-21]。另外,敲除AtNLP7后,擬南芥表現出明顯的缺氮狀態,也表現出更強的抗旱性[24]。原生質免疫共沉淀雜交技術分析也表明擬南芥AtNLP7參與到氮代謝及相關的途徑中[23,32]。本研究發現,在氮素代謝中發揮著重要功能的NLP轉錄因子的表達響應干旱脅迫,這對于解析柑橘在干旱條件下氮素調控的分子機制非常重要。
KONISHI等分析了幾種高等植物的NiR啟動子的序列,鑒定出一個保守序列(5′-tGACcCTTN10AAGagtcc-3′)作為 NRE,而且在擬南芥NiR啟動子中的NRE可以直接受硝酸鹽應答轉錄,但突變了該保守序列后,啟動子的硝酸鹽響應活性顯著降低,表明NRE是硝酸鹽應答轉錄所必需的[33-34]。通過序列分析,在PtNiR啟動子區域-196至-154的位置發現了疑似 NRE,并且應用酵母單雜交分析確定了 PtNLP2、PtNLP4、PtNLP7及PtNLP8轉錄因子均能與4×NRE位點結合互作,激活下游基因的表達。這進一步在理論上表明NLP可以通過綁定NRE位點激活PtNiR的表達。因此,枳NLP轉錄因子在柑橘的氮素代謝中可能也起著非常重要的作用,其表達水平不僅影響柑橘對氮素的吸收與同化,也與柑橘的抗旱性有著某種聯系。
4 結論
干旱脅迫影響枳的全氮含量以及PtNiR和NLP基因的表達,特別是側根中全氮含量顯著下降,其PtNiR表達受到干旱脅迫的抑制,且隨著干旱時間的延長其表達量顯著下調。PtNLP2、PtNLP4、PtNLP7及PtNLP8的表達均響應干旱脅迫但存在差異。在PtNiR啟動子中發現了硝酸鹽響應順式作用元件(NRE),PtNLP2、PtNLP4、PtNLP7及PtNLP8轉錄因子均能與NRE位點綁定互作,從而激活下游基因的表達。
