流感病毒PA蛋白與宿主蛋白PCBP1的相互作用
2018-09-11 06:19:20趙青青李俊平梁立濱黃山雨周陳陳趙玉輝王倩周圓姜麗陳化蘭李呈軍
中國農業科學 2018年17期
趙青青,李俊平,梁立濱,黃山雨,周陳陳,趙玉輝,王倩,周圓,姜麗,陳化蘭,李呈軍
(中國農業科學院哈爾濱獸醫研究所/獸醫生物技術國家重點實驗室/農業部動物流感重點開放實驗室,哈爾濱 150069)
0 引言
【研究意義】流感病毒(influenza virus)是引起流行性感冒的重要病原體,在人群中傳播可造成季節性流感以及流感大流行,嚴重危害人類的健康[1-3]。目前對于流感病毒的防控措施主要有疫苗免疫和抗流感藥物。由于流感病毒基因組包含 8條單股負鏈 RNA片段,極易發生重組和突變[4-6],使得疫苗和抗流感藥物失去保護作用。因此,研究流感病毒在宿主體內的復制及調控機制可以為防控流感病毒提供新思路。在病毒入侵宿主過程中,病毒組分與宿主蛋白發生廣泛且復雜的相互作用。PA蛋白是流感病毒RNA聚合酶復合體的重要組成部分,在流感病毒 RNA的轉錄復制過程中發揮重要作用[7]。因此研究 PA蛋白與宿主蛋白的相互作用對于揭示流感病毒在宿主體內的復制及調控機制具有重要意義。【前人研究進展】PA蛋白由流感病毒基因組第3節段編碼,全長為716個氨基酸,具有核定位信號[8]。在流感病毒粒子中,PA與PB2和PB1組成RNA聚合酶復合體,負責病毒基因組的轉錄和復制[9-10]。PA蛋白經胰酶切割后可分為兩個結構域,即氨基端結構域和羧基端結構域[11],氨基端結構域具有切割宿主mRNA前體分子5′帽子結構的內切核酸酶活性[12],羧基端的突變可使 RNA聚合酶復合體失去轉錄活性[13]。此外,PA蛋白具有蛋白水解活性[14],還是酪氨酸激酶II的底物,可以發生絲氨酸和蘇氨酸的磷酸化[15]。已發現與PA蛋白相互作用的蛋白有:IRF3[16]、MCM[17]、COPI[17]、HAX1[18]、CLE[19-20]、FACT[19]等。【本研究切入點】鑒于PA蛋白在流感病毒 RNA轉錄復制過程中的重要功能,中國農業科學院哈爾濱獸醫研究所獸醫生物技術國家重點實驗室前期利用酵母雙雜交技術從cDNA文庫中篩選與PA蛋白相互作用的宿主蛋白,獲得多聚胞嘧啶結合蛋白1(poly(rC)-binding protein 1,PCBP1)。已有研究表明,PCBP1蛋白與豬繁殖障礙呼吸綜合征病毒(PRRSV)[21]、豬瘟病毒(CSFV)[22]以及人類免疫缺陷性病毒(HIV)[23]的復制有關,且PCBP1蛋白對不同病毒復制的調控作用具有多樣性,目前尚未有關于PCBP1蛋白對流感病毒復制影響的研究。【擬解決的關鍵問題】探索PA蛋白與PCBP1蛋白的相互作用關系以及PCBP1蛋白對流感病毒復制的影響,為深入理解流感病毒在宿主體內的復制及調控機制提供數據。
1 材料與方法
試驗于 2017年在中國農業科學院哈爾濱獸醫研究所完成。
1.1 主要試驗材料
真核表達載體 pCAGGS由美國威斯康星大學Yoshihiro Kawaoka教授惠贈; pGADT7-DEST酵母雙雜交文庫由Invitrogen構建;siRNA由蘇州吉瑪基因公司合成;Flag、Myc等標簽抗體均購自Sigma-Aldrich;兔抗 PA多抗為中國農業科學院哈爾濱獸醫研究所獸醫生物技術國家重點實驗室制備;小鼠抗PCBP1單抗購自Abcam;IRDye800CW山羊抗小鼠和山羊抗兔IgG購自LI-COR Bioscience;流感病毒 A/Anhui/2/2005(H5N1)、A/WSN/1933(H1N1)均由中國農業科學院哈爾濱獸醫研究所獸醫生物技術國家重點實驗室保存。
1.2 酵母回交驗證
采用LiAc方法[24]將陰性對照組pGBKT7-Lamin+pGADT7-T、陽性對照組pGBKT7-p53+pGADT7-T和誘餌質粒 pGBKT7-PA+重組陽性質粒 pGADT7-PCBP1三組質粒分別共轉化Y2H Gold酵母感受態細胞,把轉化后的菌液分別涂布在 SD/-2、SD/-4和SD/-4/X/A三種營養缺陷平板上,30℃培養箱倒置培養5—7 d,觀察菌落生長情況及顏色。
1.3 真核表達質粒的構建及鑒定
參照GenBank中登錄的A/Anhui/2/2005(H5N1)病毒 PA基因序列(GenBank: HM172310.1)及人源PCBP1 序列(poly(rC)binding protein 1 [Homo sapiens(human)] Gene ID: 5093),設計相關克隆引物(表1)。Trizol法[25-26]提取流感病毒RNA,經12 bp通用引物反轉錄得到cDNA,利用PA特異性擴增引物,從流感病毒cDNA中擴增出PA片段,與NdeI/SmaI雙酶切處理的pCAGGS載體連接,構建重組表達質粒pCAGGS-Flag-PA;根據QIAGEN的RNA提取試劑盒說明書提取A549細胞總RNA,以oligo(d)T為引物反轉錄制備cDNA,利用PCBP1特異性擴增引物擴增 PCBP1目的基因,將載體 pCAGGS進行EcoRI/SmaI雙酶切處理,構建表達質粒 pCAGGSMyc-PCBP1。

表1 本研究所使用的引物Table 1 Primers used in this study
1.4 免疫共沉淀試驗
將HEK293T細胞按1∶4傳代于賴氨酸包被的6孔板中,當細胞密度達到約 80%時,參照轉染試劑Lipofectamine LTX with Plus Reagent(Invitrogen)說明書,將真核表達質粒pCAGGS-Flag-PA和pCAGGSMyc-PCBP1各1 μg配制單轉和共轉混合液,室溫靜置25 min,然后將HEK293T細胞用PBS洗一遍后每孔加入500 μL opti-MEM培養基,然后加入相應的轉染混合液,8—12 h后更換正常生長培養基。轉染48 h后,每孔加入 200 μL含 Protease Inhibitor Cocktail(Roche)的NP40裂解液,在冰上裂解細胞30 min,4℃ 12 000 r/min離心15 min并收集上清,取1/10樣品留作裂解液對照Input,剩余樣品經加入1μL小鼠抗FLAG 單抗 4℃結合4—6 h、加入30 μL Protein G瓊脂糖珠(Roche)4℃結合 4—6 h、預冷裂解液(含PMSF)洗4—6次、加入30 μL 2×蛋白上樣緩沖液后95℃煮5 min等一系列過程制備IP樣品。將得到的Input和IP樣品進行SDS-PAGE電泳、300 mA恒流轉膜2 h、5%脫脂乳室溫封閉1 h、兔抗Myc多抗和兔抗 FLAG多抗(1∶1 000稀釋)室溫孵育 1 h、0.05%PBST洗3遍、IRDye800CW山羊抗兔IgG 二抗(1∶10 000稀釋)室溫孵育1 h、0.05%PBST洗3遍,然后利用Odyssey雙色紅外熒光成像系統(LI-COR公司)掃膜。
1.5 流感病毒蝕斑滴定
將MDCK細胞接種于12孔板,當細胞密度達到95%時進行蝕斑滴定。配制2×MEM培養基(含0.6%BSA,1 μg·mL-1TPCK 胰酶),用等體積 ddH2O 將2×MEM 培養基稀釋為 1×MEM 培養基,并分裝于1.5 mL離心管,每管900 μL,取100 μL病毒液進行倍比稀釋。將12孔板中的MDCK細胞用PBS洗兩遍,然后每孔加入100 μL倍比稀釋后的病毒液,感染1 h。然后棄病毒液,每孔加入1 mL固態培養基(2×MEM培養基、等體積的 2%低熔點瓊脂糖凝膠),待培養基凝固后,倒置于CO2培養箱繼續培養,48 h后用福爾馬林固定液固定。固定后,棄孔中固態培養基,以0.1%結晶紫溶液染色,空斑計數。應用GraphPad Prism 6.0進行Multiple t tests分析。
1.6 PCBP1蛋白過表達對病毒復制的影響
參照慢病毒包裝系統 pLVX-IRES-ZsGreen1(Clontech)說明書,將pLVX-IRES-ZsGreen1空載體或 pLVX-IRES-ZsGreen1-PCBP1、pSPAX2和 pMDG按4∶3∶1共同轉染HEK293T細胞包裝假病毒,48 h后收獲上清,將包裝的病毒侵染A549細胞,被假病毒侵染的A549細胞將表達綠色熒光蛋白ZsGreen1,48 h后將A549細胞消化成單細胞根據熒光標記進行超速流式細胞分選,留下陽性細胞繼續培養,建立PCBP1蛋白過表達A549的細胞系,并通過Western blot驗證PCBP1蛋白的過表達情況。將A/WSN/1933(H1N1)毒株以MOI=0.01感染對照組及PCBP1蛋白過表達A549細胞系,并分別于感染后24、48 h收獲上清,進行蝕斑滴定計數。
1.7 PCBP1基因表達下調對病毒復制的影響
參照Lipofectamine RNAiMAX(Invitrogen)說明書,將 Scramble siRNA和 PCBP1 siRNA分別轉染A549細胞,轉染后48 h以MOI=0.01感染A/WSN/1933(H1N1),并于感染后24、48 h收上清進行蝕斑滴定計數,同時收獲細胞樣品進行Western blot檢測干擾效果。
2 結果
2.1 酵母回交驗證
將陰性對照組、陽性對照組和 pGADT7-PCBP1+pGBKT7-PA 3組酵母質粒分別共轉化Y2H感受態細胞后涂在SD/-2、SD/-4和SD/-4/X/A 3種營養缺陷平板上。結果顯示,陰性對照組 pGBKT7-Lamin+pGADT7-T可以在SD/-2平板上生長,但不能在SD/-4、SD/-4/X/A平板上生長,陽性對照組 pGBKT7-p53+pGADT7-T在3種缺陷型平板上均能正常生長,而且陽性對照組可以分解SD/-4/X/A平板的X-α-Gal,使菌落變藍,表明陰性對照和陽性對照均正常。重組陽性質粒pGADT7-PCBP1+pGBKT7-PA也可以在SD/-2、SD/-4、SD/-4/X/A 3種營養缺陷型培養基上正常生長,并且也能分解底物X-α-Gal,使菌落呈現藍色(圖1),與陽性對照組一致,表明PA蛋白與PCBP1蛋白在酵母系統中存在相互作用。
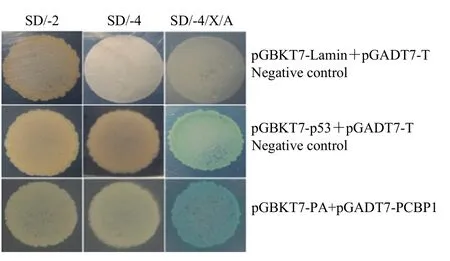
圖1 酵母回交驗證PA蛋白與PCBP1蛋白的相互作用Fig. 1 Interaction between PA protein and PCBP1 protein in yeast
2.2 PA蛋白與PCBP1蛋白互作的Co-IP驗證
將pCAGGS-Flag-PA和pCAGGS-Myc-PCBP1共轉染 HEK293T細胞。細胞裂解液樣品(Input)結果顯示, PA蛋白和PCBP1蛋白均正常表達,Co-IP結果顯示PA蛋白與PCBP1蛋白之間存在相互作用,而PA蛋白與pCAGGS空載體不存在相互作用(圖2)。
2.3 過表達PCBP1蛋白抑制流感病毒復制
利用慢病毒載體pLVX-IRES-ZsGreen1包裝假病毒,侵染A549細胞后經流式分選構建PCBP1蛋白過表達細胞系。取部分細胞樣品進行Western blot檢測,證實PCBP1蛋白在A549過表達細胞系中表達量顯著上調(圖3-A)。同時,將A/WSN/1933(H1N1)毒株以MOI=0.01感染A549過表達細胞系和對照細胞系,收上清后進行蝕斑滴定檢測病毒復制情況。結果顯示,感染后48 h病毒滴度下降,P<0.01,差異極顯著,說明PCBP1蛋白過表達后可以抑制流感病毒的復制(圖3-B)。
2.4 PCBP1蛋白表達下調后促進流感病毒的復制
將 Scramble siRNA和 PCBP1 siRNA分別轉染A549細胞后,進行Western blot檢測,確證PCBP1蛋白表達水平下降(圖 4-A)。同時,以 MOI=0.01 A/WSN/1933(H1N1)病毒感染后,利用蝕斑試驗檢測病毒復制情況。結果顯示,感染后48 h病毒滴度升高,P<0.01,差異極顯著(圖 4-B),表明 PCBP1蛋白表達下調后促進病毒復制。
3 討論
酵母雙雜交技術是高通量篩選和鑒定蛋白互作的有效方法[27],廣泛應用于篩選與病毒蛋白相互作用的宿主蛋白。由于流感病毒基因組較小,蛋白編碼能力有限,因此在病毒感染宿主的過程中會與大量的宿主蛋白發生相互作用。筆者前期利用酵母雙雜交技術從人源細胞cDNA文庫中篩選到與流感病毒PA蛋白相互作用的宿主蛋白PCBP1。PCBP1是一種RNA結合蛋白,因其可以與 RNA多聚胞嘧啶區域特異性結合而得名。PCBP1基因在1994年由AASHEIM等從人淋巴細胞cDNA文庫中克隆出來[28],屬于多聚胞嘧啶結合蛋白PCBPs家族成員之一,因其含有3個與該家族最先被發現的成員hnRNP K(heterogenous nuclear ribonucleoprotein K)相似的 KH結構域(hnRNP K homology, KH domain),因此又被命名為 hnRNP E1[29]。PCBP1蛋白廣泛表達于人體各組織器官,在多水平參與基因的表達調控,如轉錄、前體mRNA剪切、mRNA穩定性、翻譯沉默或翻譯增強及蛋白質之間相互作用等[29-30]。PCBP1含有核定位信號[31],具有核-質穿梭特性[32]。有研究表明,PCBP1及PCBP2可與豬繁殖障礙呼吸綜合征病毒(PRRSV)的NS 1β蛋白互作,促進PRRSV的復制[21];PCBP1與豬瘟病毒(CSFV)的Npro蛋白互作,PCBP1對豬瘟病毒具有正調控作用[22]。在人類免疫缺陷性病毒HIV的研究中發現,PCBP1可以同HIV的外顯子剪接抑制子3(exon splicing silencer 3,ESS3)區域相互作用,且對病毒復制起負調控作用[23]。可見,PCBP1蛋白對不同病毒復制的調控作用具有多樣性。目前也有許多研究報道PCBP1與人類多種腫瘤發生如結腸癌、乳腺癌及宮頸癌等具有相關性[33-35]。PCBP1功能的多樣性可能是由于其參與不同的通路或形成不同的復合體,從而對mRNA穩定性及蛋白翻譯過程影響不同。

圖2 Co-IP 驗證PA蛋白與PCBP1蛋白相互作用Fig. 2 Verification of interaction between PA protein and PCBP1 protein by Co-IP assay

圖3 過表達PCBP1 抑制流感病毒復制Fig. 3 Inhibition of influenza virus replication in PCBP1-overexpressing A549 cells

圖4 敲低PCBP1促進流感病毒復制Fig. 4 Knockdown of PCBP1 in A549 cells promotes influenza virus replication
本研究通過酵母回交驗證試驗證實了 PCBP1蛋白與流感病毒PA蛋白在酵母系統中存在相互作用。由于酵母雙雜交系統假陽性率較高,且蛋白質在酵母與哺乳動物細胞中的加工修飾不完全相同,因而蛋白質在酵母中的相互作用并不一定反應真實情況。本研究采用免疫共沉淀檢測流感病毒PA蛋白與PCBP1蛋白在HEK293T細胞中相互作用,發現PA蛋白可以與PCBP1蛋白發生免疫共沉淀,說明PA蛋白與PCBP1蛋白在哺乳動物細胞中存在相互作用。本研究發現利用慢病毒過表達系統 pLVX-IRES-ZsGreen1將 A549細胞中PCBP1蛋白過表達,可以抑制流感病毒復制,而通過 siRNA干擾下調A549細胞中PCBP1蛋白表達后,可以促進流感病毒復制,即PCBP1蛋白對流感病毒復制具有負調控作用,但其調控流感病毒復制的具體機制仍有待進一步研究。
4 結論
本研究發現流感病毒PA蛋白與宿主蛋白PCBP1在酵母和哺乳動物細胞中存在相互作用,且宿主蛋白PCBP1對流感病毒復制具有負調控作用。
