髓系來源的抑制細胞在肝病發病中的作用研究進展*
2018-09-20 05:59:18高苗苗張紀元綜述鄒正升審校
實用肝臟病雜志 2018年5期
關鍵詞:研究
高苗苗,張紀元 綜述,鄒正升 審校
2013年全球疾病負擔研究表明,我國居民所有死亡原因中,肝臟疾病所致死亡的比例為5.98%,給社會造成極大的經濟負擔。機體的免疫功能在肝病的進展中發揮重要作用,當前的研究也表明多種免疫細胞參與調節肝病的進展。髓系來源抑制細胞(Myeloid derived suppressor cells,MDSC)是一群異質性細胞。正常生理情況下,髓系祖細胞可分化為成熟的粒細胞、巨噬細胞、樹突狀細胞(Dendritic cell,DC)。然而在腫瘤、感染、敗血癥、外傷等病理狀態下,髓系祖細胞及未成熟髓系細胞的成熟受阻,停留在分化的各個階段,成為MDSC[1,2]。MDSC具有廣泛的調節作用,在多種疾病的進展中發揮著重要作用。近年來,MDSC在免疫調節方面發揮的作用受到越來越多的關注。
1 MDSC的歷史與亞群
1978年,一項關于卡介苗對脾臟免疫影響的研究中發現脾臟中存在抑制免疫的細胞,實驗證明這些抑制性細胞來自骨髓。由于這些異常的髓系細胞缺乏成熟T細胞、B細胞、自然殺傷細胞(Natural killer cell,NK)及巨噬細胞的膜性標記物,因此被命名為失效細胞、否決細胞或者自然抑制細胞[3]。隨后的多項研究證實這些細胞在腫瘤等疾病中擴增,并能夠抑制淋巴細胞數量、細胞毒性T細胞誘導和激活、以及DC細胞的活化等[4]。研究發現 GM-CSF[5]、G-CSF[6]、VEGF、iNOS[7]、STAT3、MyD88 和 NF-κB[8]、Flt3L、SCF 和 S100A8/9、趨化因子[9]、miRNA-494、ASAM10和Notch等促使這些異常的髓系細胞擴增,相反,全反式維甲酸、Sutent等抑制其增長。直到2007年,Gabrilovich DI等發表文章,將此類細胞命名為MDSC達成一致[10]。
小鼠MDSC的共同特點是均可檢測到其細胞表面表達GR1和CD11b分子。GR1包括Ly6C和Ly6G,分別表達于巨噬細胞和中性粒細胞。CD11b是巨噬細胞的標記物。根據Ly-6C與Ly-6G的表達,小鼠MDSC細胞分為兩類,CD11b+Ly-6GlowLy-6Chigh細胞是類單核細胞形態,主要表達氮氧合酶,T細胞抑制性活動增強,定義為單核細胞型MDSC(mMDSC)。與此相對,CD11b+Ly-6G+Ly-6Clow 細胞是類粒細胞形態,高表達精氨酸酶 -1(Arginase 1,Arg-1),定義為粒細胞型MDSC(gMDSC)。后者有多核形態,因此,多核形態的MDSC也稱作gMDSC[11]。另外一些研究發現,MDSC的某些亞群表達IL-4和IL-13(IL-4Rα)受體的α鏈,另一種巨噬細胞標記(F4/80),M-CSF-1R 或 c-fms(CD115),和共刺激分子CD80。
與小鼠的MDSC類似,人的MDSC也可分為mMDSC與gMDSC兩群。mMDSC表型為 CD11b+CD33+HLA-DR-C D14+CD15-,相對的,gMDSC 為 CD11b+CD33+HLA-DR-CD14-CD15+[3]。隨著研究的進一步深入,MDSC更多的表型被發現,包括高表達的CD66b,低表達的CD62L和CD16[12],血管內皮細胞生長因子受體1[13]及S100A9[14]。
2 MDSC的作用機制
多種證據表明,MDSC參與維持機體的免疫耐受,通過多種免疫學機制調節人體的固有免疫和適應性免疫。
精氨酸是T細胞活化的必需氨基酸,MDSC抑制CD4+和CD8+T細胞攝取精氨酸,同時其細胞內高水平的精氨酸酶-1耗盡環境中的精氨酸[15]。MDSC產生的活性氧和過氧化亞硝酸鹽通過催化TCR的硝基化抑制CD8+T細胞,從而防止T細胞抗原肽的相互作用[16]。腫瘤浸潤可能刺激MDSC產生 TGF-β,從而誘導調節性 T 細胞(Regulatory cell,Treg)產生,間接抑制T細胞活化[17]。MDSC還可下調TCR相關的ζ-鏈,后者是CD4+/CD8+T細胞活化所需信號。MDSC下調CD62L(一種幼稚T細胞歸巢到淋巴結必要的膜分子),使得T細胞無法遷移到淋巴結。MDSC還可通過剝奪半胱氨酸抑制T細胞活化。半胱氨酸是T細胞活化的另外一種必需氨基酸。T細胞的半胱氨酸來自DC細胞和巨噬細胞,它們利用蛋氨酸和細胞外胱氨酸合成半胱氨酸。當MDSC高濃度聚集時,吸收大部分可用的胱氨酸,使得DC細胞和巨噬細胞合成半胱氨酸受阻。由于得不到足量的半胱氨酸,T細胞合成激活相關的蛋白受阻[18]。
MDSC也通過產生IL-10、TGF-β或精氨酸酶-1誘導Treg,間接影響T細胞活化,進而下調細胞免疫[19]。MDSC還可使得抗腫瘤免疫應答轉變為促進腫瘤發展的2型應答,其產生的細胞因子IL-10可下調巨噬細胞產生的IL-12。另一方面,巨噬細胞可以反作用于MDSC,促進其產生更多的IL-10,進一步放大2型促腫瘤免疫應答。雖然MDSC調節NK細胞的作用尚不明確,但一些研究表明,MDSC可以抑制NK細胞對腫瘤細胞的細胞毒作用并下調NK細胞產生IFN-γ,進一步的研究發現NKG2D是NK細胞活化所需的一種受體,MDSC的這種抑制NK細胞功能的作用就是通過阻斷NKG2D的表達[20]。然而,另一項研究表明,MDSC表達NKG2D配體Rae-1,從而激活NK細胞[21],提示激活的NK細胞也可以殺死MDSC。這些研究結果的差異可能源于MDSC不同亞群的差異。
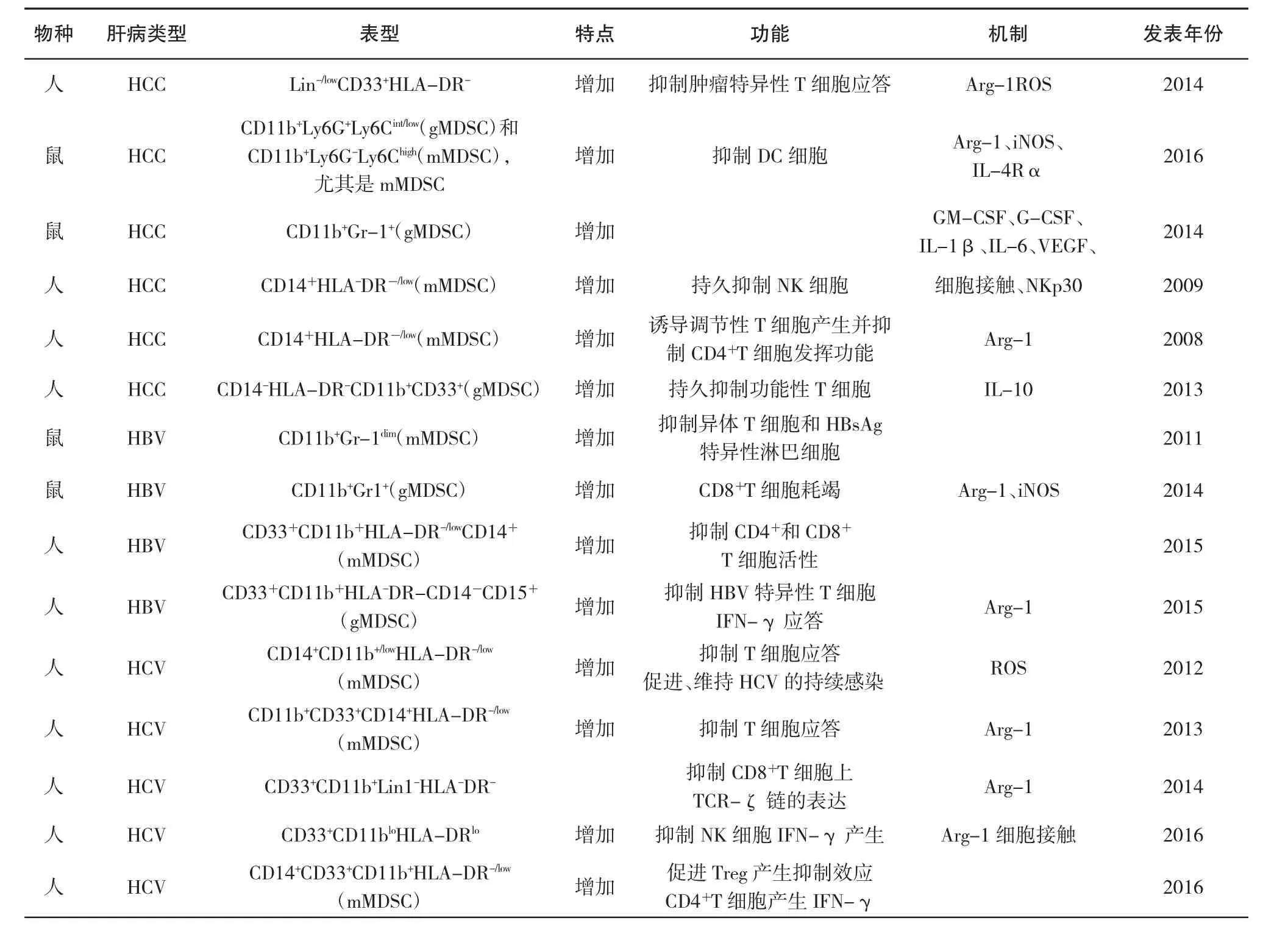
表1 MDSC在人和小鼠肝臟疾病中的調節作用
3 MDSC與肝臟疾病
近年研究發現,MDSC與肝臟疾病的發生發展具有密切關系,通過多種機制作用于肝臟疾病的發生發展。近年來,MDSC在人肝臟疾病中發揮的作用越來越受到重視,現就肝臟疾病中的MDSC研究現狀做一綜述。表1為近年來關于MDSC在人和小鼠肝臟疾病中發揮調節作用的相關研究及其作用機制的匯總。
3.1 MDSC與肝癌MDSC能夠有效抑制抗腫瘤免疫 Shen研究發現,未治療的HCC患者外周血Lin-/lowCD33+HLA-DRMDSC細胞頻率明顯高于健康對照組,升高的頻率與癌癥嚴重程度呈正相關,三四期肝癌患者MDSC細胞頻率顯著高于一二期患者,而后者顯著高于健康對照組。有趣的是,研究發現伴有脾腫大的肝癌患者與不伴有者相比,其MDSC水平頻率進一步升高。MDSC對T細胞增殖及IFN-γ水平的抑制程度與癌癥的嚴重程度呈正相關,隨著癌癥分期的增加,其IFN-γ濃度逐級降低。晚期肝癌患者MMP13表達增加,與MDSC水平呈正相關;而其COX-2表達減少,與MDSC水平呈負相關[22]。Xu研究表明,活化的肝星狀細胞能夠誘導骨髓來源細胞分化為MDSC,其中對mMDSC的產生影響最顯著。肝星狀細胞通過精氨酸酶-1、iNOS、IL-4Rα抑制DC細胞形成并促進MDSC的生成。肝星狀細胞還可通過COX2-PGE2-EP4信號通路促進MDSC的產生。肝星狀細胞產生PGE2,作用于MDSC表面的EP4受體。因而EP4受體拮抗劑可阻斷MDSC的積累。同樣的,使用COX-2阻斷劑可抑制HSC誘導MDSC的產生。并發現該通路主要作用于gMDSC,對 mMDSC 的作用微乎其微[23]。Kapanadze T et al研究發現,腫瘤誘導產生的GM-CSF介導肝癌MDSC的累積[24]。γδ T細胞來源的IL-17具有同樣的作用。IL-17誘導腫瘤分泌CXCL5,然后通過CXCR2參與招募MDSC。此外,IL-17也直接作用于MDSC,提高它們的抑制能力。反過來,MDSC通過釋放 IL-23和 IL-1β增加 γδT細胞產生 IL-17[25]。Hoechst等研究發現,在肝癌患者,MDSC對NK細胞的抑制作用主要是通過細胞接觸的方式下調NKp30,而與精氨酸酶-1及iNOS通路無關[26]。在肝癌患者的體外實驗中,CD14+HLA-DR-/low誘導與CD4+T細胞共培養的CD4+CD25+Foxp3+調節性T細胞,從而發揮其抑制作用[27]。Fumitaka研究發現,MDSC頻率與肝癌的進展有關,而與其纖維化和炎癥程度無關。并且患者經過射頻消融等治療后的療效與體內MDSC的頻率呈負相關,這些研究提示抑制MDSC頻率可能會改善患者預后[28]。
3.2 MDSC與乙型肝炎 Chen發現,在HBV轉基因小鼠,肝臟CD11b+Gr-1-MDSC頻次大約是正常小鼠肝臟的兩倍。HBV轉基因小鼠的肝臟源性的CD11b+Gr-1-MDSC也抑制異體T細胞和HBsAg特異性淋巴細胞的增殖能力[29]。Kong研究發現,HBV持續感染的小鼠,其γδT細胞能夠誘導CD11b+Gr1+MDSC 介導的CD8+T細胞耗竭[30]。Anthony研究表明,在HIV患者中,高頻的MDSC有助于第16周HBV疫苗的應答。Lu研究發現,慢性乙型肝炎患者外周血Lin-HLA-DR-CD11b+CD33+MDSC頻率顯著高于健康人,且其頻率在HBeAg+活動期與ALT、AST及TBil呈負相關,而與ALB呈正相關。Fang發現,慢性乙型肝炎患者血中mMDSC的水平與血清中HBsAg的高濃度呈正相關,而急性乙型肝炎患者的低水平HBsAg可能不會誘導mMDSC的積聚。研究發現,HBsAg以IL-6依賴的方式誘導STAT3的磷酸化,IL-6與單核細胞上的受體結合通常會激活轉錄因子STAT3。另一方面,HBsAg激活ERK1/2信號通路,從而誘導IL-6和MDSCs的分化。與此不同的是,Laura等研究發現,慢性乙型肝炎患者的mMDSC頻率與健康對照組無顯著差別,而其體內的gMDSC的頻率是顯著增加的。研究雖然沒有發現gMDSC數量與病毒載量、HBsAg及HBeAg等之間有相關性,但在慢性乙型肝炎疾病進展的四個分期中,處于免疫耐受期(即HBV持續復制而不伴有肝臟炎癥)的患者,其gMDSC的頻率明顯高于其他期,這一研究提示gMDSC可能參與維持乙型肝炎的免疫耐受。慢性乙型肝炎患者gMDSC高表達精氨酸酶-1,并且慢性乙型肝炎患者循環血中的精氨酸酶+gMDSC比例明顯高于健康對照組,其肝內精氨酸酶+gMDSC富集,高于循環血中的頻率。研究顯示,gMDSC產生大量精氨酸酶消耗精氨酸,抑制T細胞功能,進而損傷肝臟。
3.3 MDSC與丙型肝炎 Tacke發現,HCV可以提高CD33+CD14+CD11b+/lowHLA-DR-/lowMDSC表達,并上調ROS介導抑制T細胞反應,進而促進、維持HCV的持續感染。Cai發現,慢性丙型肝炎患者的MDSC水平與血漿HCV-RNA,ALT,AST,活化的CD38+T細胞水平呈正相關,并以精氨酸-1-依賴途徑抑制T細胞功能。此外還發現,慢性丙型肝炎患者進行臨床治療后,MDSC水平最早在第4周時開始下降,但在治療12周后反彈。Zeng研究發現,慢性丙型肝炎患者外周血以及肝內CD8+T細胞表面的ζ鏈表達均下調,并與未治療的慢性丙型肝炎患者MDSC頻率呈負相關。Pang研究表明,HCV核心蛋白通過上調吲哚胺2,3-雙加氧酶表達及抑制T細胞增殖促進CD14+單核細胞轉變為CD14+HLA-DR/低表型(即mMDSC)。更重要的是,HCV通過PI3K通路增加IL-10和TNF-α分泌,從而誘導mMDSC擴增。Goh研究發現,與未感染HCV的CD33+PBMCs相比,NK細胞與HCV感染的CD33+PBMCs共培養產生IFN-γ較少。而兩組的NK細胞表達量并無差別,因此,MDSC下調NK細胞產生IFN-γ可能發生在轉錄后。并發現了CD33+CD11blowHLA-DRlowMDSC通過精氨酸酶-1耗竭精氨酸,進而抑制mTOR通路介導的細胞增殖以及細胞因子的產生,例如抑制NK細胞產生IFN-γ。Ren研究顯示,HCV誘導mMDSC高表達磷酸化信號轉導與轉錄激活因子3和IL-10,而阻斷STAT3信號通路會抑制HCV介導的mMDSC的擴增及IL-10的產生。同時發現,HCV誘導的MDSC可以顯著增加健康人來源的PBMC中CD4+CD25+Foxp3+調節性T細胞的數量,而慢性丙型肝炎患者的MDSC減少導致其Foxp3+Treg細胞顯著減少以及效應CD4+T細胞產生的IFN-γ顯著增加。這些結果表明,HCV誘導的MDSC促進Treg細胞的增加和抑制效應CD4+T細胞的功能。
4 小結與展望
目前為止,關于MDSC與肝臟疾病的研究取得了很大的進步,多項研究表明MDSC在肝臟的疾病狀態下表達上調,通過精氨酸酶、誘導型一氧化氮合酶等途徑抑制T細胞應答、促進T細胞耗竭,且在肝癌中誘導調節性T細胞產生;通過細胞接觸和NKp30途徑持久抑制NK細胞的功能;通過IL-4Rα及精氨酸酶等途徑抑制DC細胞功能,經多種途徑調節肝臟的免疫狀態,在肝臟疾病的發生發展中發揮著重要的作用。因此調節MDSC及其相關分子信號有可能成為治療肝病的新靶點。值得注意的是,當前關于MDSC在肝臟疾病中的研究主要集中在肝癌、乙型肝炎和丙型肝炎,隨著乙型肝炎疫苗的廣泛使用、輸血管理的加強和有效抗病毒藥物的使用,我國乙型肝炎、丙型肝炎的感染率逐年下降。而伴隨著人民生活水平的不斷提高,嗜酒人群比例不斷上升,酒精相關的健康問題逐漸凸顯,不容忽視。一項多中心的研究表明,2000~2004年肝病住院患者中酒精性肝病患者分別占2.7%、2.9%、3.0%、3.6%和4.4%,所占比例呈不斷上升趨勢,5年間上升了1.63倍。而目前為止,MDSC與酒精性肝病的發生發展的關系尚缺乏研究,因此闡示MDSC在酒精性肝病中的免疫學特點,一方面可以彌補這一空白,另一方面有可能發現酒精性肝病新的治療靶點。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
遼金歷史與考古(2019年0期)2020-01-06 07:45:20
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
電子制作(2018年11期)2018-08-04 03:26:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
國際商務財會(2017年8期)2017-06-21 06:14:14
電子制作(2017年23期)2017-02-02 07:17:19
