馬尾松林土壤微生群落結構對不同營林處理的響應
2018-09-28 08:45:04肖文發曾立雄黃志霖高尚坤張維誠
生態學報 2018年16期
關鍵詞:結構
雷 蕾,肖文發,曾立雄,*,黃志霖,高尚坤, 張維誠,王 松
1 中國林業科學研究院森林生態環境與保護研究所,國家林業局森林生態環境重點實驗室,北京 100091 2 南京林業大學南方現代林業協同創新中心,南京 210037 3 華南農業大學林業風景園林學院,廣州 510642 4 湖北省秭歸縣國有九嶺頭林場,宜昌 443604
多樣的森林類型和不同營林措施的干擾造成了森林土壤碳庫維持機制、碳固定過程研究較大的不確定性[1]。土壤微生物是土壤中物質轉化的動力,對土壤有機質的分解起著重要作用[2- 4],且對環境變化十分敏感[5]。森林采伐等對森林生態系統產生干擾的經營措施極可能會通過改變地上植被組成、凋落物和根系分泌物等有機質碳輸入的數量和化學成分,影響微生物的底物獲取和微生物過程[6- 7],從而作用于土壤有機碳的分解與轉化,引起土壤碳排放的變異[2, 8- 9]。近些年來許多學者針對森林經營特別是采伐措施對土壤碳排放的影響進行了大量研究[10- 12],但對于采伐干擾后微生物的響應研究有限[13],深入分析森林采伐后微生物過程的變化是探討森林采伐對于土壤碳庫的影響機制的關鍵部分,是評估采伐干擾對森林土壤碳庫過程的影響機制的重點與難點[12]。
馬尾松(Pinusmassoniana)是中國南方主要造林樹種,是三峽庫區分布面積最大的森林類型[14- 16],但由于本區域馬尾松林經營粗放且人為干擾強烈,低產林大面積存在,這可能是本區域森林生態系統碳密度偏低的主要可能原因。因此,在三峽庫區開展馬尾松林的經營試驗,并對經營后馬尾松林的土壤碳庫動態、碳固定過程的微生物因子進行長期連續的監測和調查十分必要。本研究以三峽庫區人為管理后馬尾松飛播林為研究對象,對營林處理后(對照(未采伐),除灌(清除灌叢),采伐1(強度15%)和采伐2(強度70%))不同時期(處理后初期(2013年12月)和處理后15個月(2014年12月))的土壤微生物生物量、微生物群落結構組成以及土壤溫濕度等環境因子進行了測定。我們假設認為地上植被組成的改變造成了土壤微生物群落結構的差異性,通過分析不同營林處理的馬尾松林土壤微生物因子的變化情況及其主要影響因子,以期為深入認識營林干擾措施對土壤碳排放過程的微生物影響機制提供科學依據。
1 研究區概況
研究地點位于湖北省秭歸縣三峽庫區九嶺頭林場,地理位置30°59′N,110°47′E,海拔156—2030.6 m,年均氣溫16.9℃,年降水量1000—1250 mm,多集中于4—9月,屬于亞熱帶大陸性季風氣候。土壤類型以黃壤、黃棕壤為主[16]。調查樣地內馬尾松林為70年代飛播造林,樣地內林木分布均勻,馬尾松為主要優勢種,蓋度為80%,伴生有少量光皮樺(Betulaluminifera)、漆樹(Toxicodendronvernicifluum)、杉木(Cunninghamialanceolata)。灌木主要有火棘(Pyracanthafortuneana)、胡枝子(Lespedezabicolor)、木姜子(Litseapungens)等,草本主要有狗脊(Woodwardiajaponica)、苔草(Carextristachya)、三脈紫菀(Asterageratoides)、中日金星蕨(Parathelypterisnipponica)等。
三峽庫區馬尾松林主要受到大樹的采伐和林下灌叢清除2種措施的干擾,但在采伐灌叢過程中,當地居民也會采伐林內新更新的光皮樺、杉木等,因此在2013年9月根據上述常見的營林干擾措施,在三峽庫區九嶺頭林場設置了3種營林干擾類型,分別為除灌(SM)措施清除樣地內所有灌木,并對灌木清理中產生的剩余物(灌叢枝葉等)進行仔細清除;采伐1(H1)措施清除樣地內徑階在4 cm以上的非馬尾松(主要為光皮樺、漆樹、杉木,包括部分高大灌木,如木姜子、火棘等),強度為15%(按胸高斷面積計算);采伐2(H2)措施清除樣地內平均胸徑(17.9 cm)以上的馬尾松,強度為70%。本實驗樣地設置于經營后的馬尾松飛播林內,樣地內采用典型采樣法,在除灌、采伐1、采伐2分別設立3塊20 m×20 m固定樣地,同時在固定樣地相鄰處,土壤條件基本相同的林地,設置對照固定樣地3塊(20 m×20 m)。伐木作業采用人力油鋸伐木,僅對采伐的樹干進行了移除,不對采伐產生的剩余物如枝葉等進行清除。各種經營措施并未對活地被層草本以及地表枯落物進行任何處理,上述營林措施于2013年10月中旬完成(樣地具體特征詳見前期研究表1[15]),且觀測期內每隔半年定期對除灌樣地內灌叢進行清除。
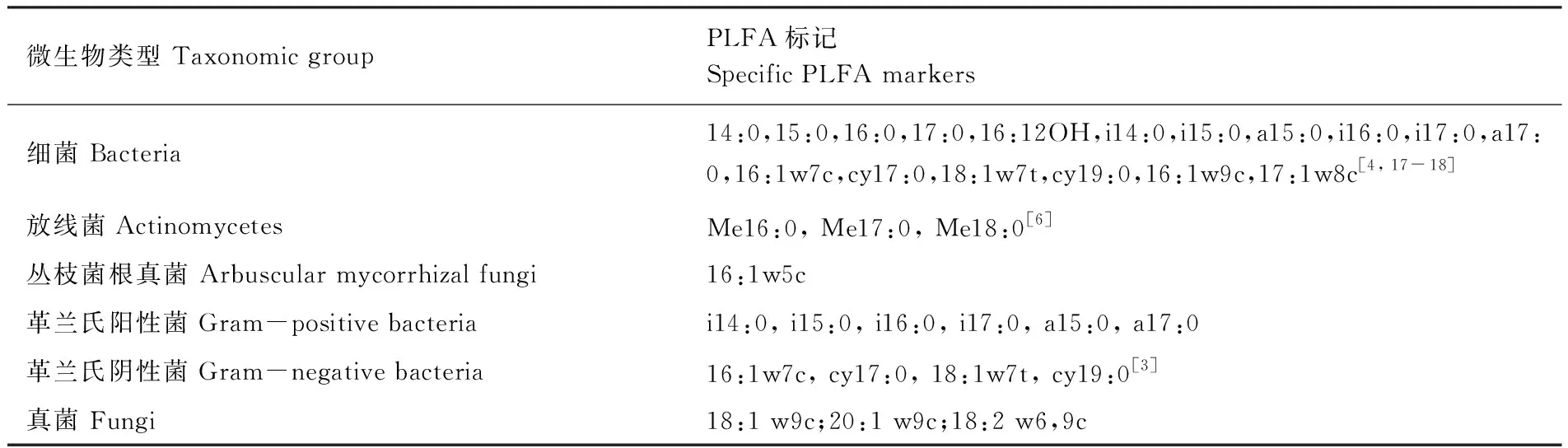
表1 表征微生物類群的PLFA
2 研究方法
2.1 土壤樣品采集與相關指標分析
在固定樣地內隨機設置3個觀測小區(1 m×1 m),于處理后初期(2013年12月),處理后15個月(2014年12月),使用土鉆對觀測小區0—10 cm土壤取3個土樣,同一固定樣地同層混合。將采集的部分新鮮土樣過2 mm篩,一部分-80℃保存,采用磷脂脂肪酸方法(phospholopid fatty acid, PLFA)對微生物群落結構(soil microbial community,SMC)進行測定和分析[17],根據不同PLFA確定微生物類群,每種類群的PLFA生物量采用nmol/g干土進行表述,表征不同類群的PLFA標記如表1所示,主要分為細菌(B)、放線菌(ACT)、叢枝菌根真菌(AMF)、革蘭氏陽性菌(GP)、革蘭氏陰性菌(GN)以及真菌(F)。一部分過篩的新鮮土樣置于4℃冰箱保存并于一周內采用氯仿熏蒸浸提法測定土壤微生物量碳(MBC)、微生物量氮(MBN)[17]。一部分過篩的新鮮土樣通過KCL溶液浸提后采用流動分析儀測定銨態氮、硝態氮。剩余樣品風干通過重鉻酸鉀外加熱法測定土壤有機碳(SOC),凱氏定氮法測定全氮(TN),pH采用玻璃電極法測定[19]。用土壤溫濕度測定儀器(Watchdog 2000 series weather stations)測定0—10 cm土壤溫度濕度。
2.2 數據統計分析
利用單因素方差分析(one-way ANOVA)檢驗不同采伐處理方式下土壤微生物生物量和土壤理化性質的差異(P<0.05),利用雙因素方差分析(two-way ANOVA)檢驗采伐處理方式和處理時間對于土壤理化指標、微生物生物量和各類微生物PLFA含量的影響。同時采用主成分分析(PCA)和冗余度分析(RDA)檢驗不同處理樣地SMC結構的差異及其與土壤理化性質的關系。主成分和冗余度分析均在軟件Canoco 4.5中進行,其余分析處理在SPSS 19.0中進行,用SigmaPlot 12.5軟件作圖。
3 結果與分析
3.1 土壤微生物生物量對不同采伐處理的響應
在干擾后初期(2013年12月),各處理間土壤微生物總PLFAs和各類微生物PLFAs差異并不顯著(圖1)。但在2014年12月,與CK相比,SM、H1與H2的放線菌呈現出降低的趨勢,且H1與H2均達到顯著水平(P<0.05)。在2014年,除AMF外,SM、H1、H2土壤微生物總生物量以及各類群生物量均呈現降低的趨勢。綜合2年數據可知,處理方式對土壤微生物量無顯著影響,而處理時間顯著影響了土壤微生物總PLFAs、放線菌PLFAs和真菌PLFAs。處理方式與處理時間對總PLFAs與各類菌群PLFAs均無顯著的交互作用(表2)。
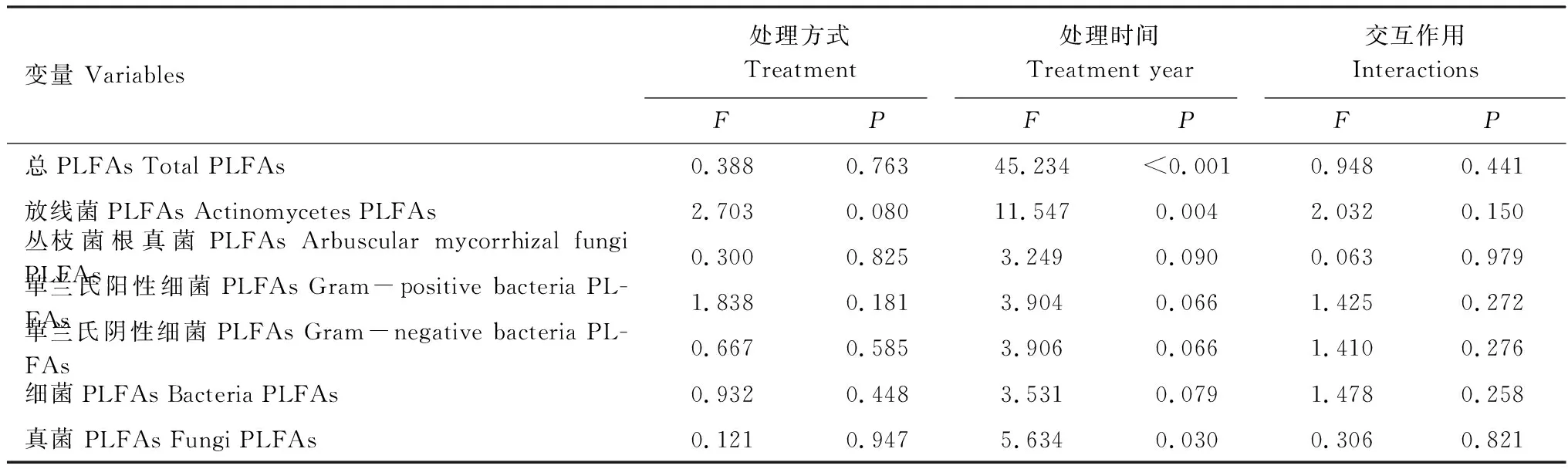
表2 處理方式、處理時間及其交互作用對土壤微生物量的影響
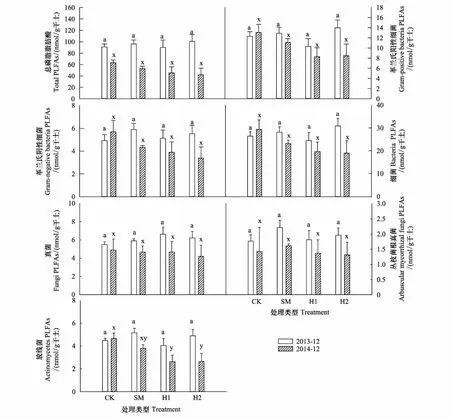
圖1 土壤微生物PLFAs對不同采伐方式的響應
3.2 土壤微生物群落結構對不同采伐處理的響應
土壤微生物群落的磷脂脂肪酸相對豐度是評價微生物群落結構變化的重要指標。對不同處理和處理時間下土壤中所提取的20種磷脂脂肪酸進行主成分分析,結果顯示,在處理后初期(2013年12月),各處理微生物群落結構并無顯著差異,第1、2主成分的貢獻率分別為60.9%、17.4%。而在2014年,CK與其他處理的微生物群落結構存在顯著差異,不同喬木層采伐處理(H1和H2)之間微生物群落差異不顯著(圖2)。

圖2 不同處理土壤微生物磷脂脂肪酸相對豐度主成分分析
通過對主成分的因子載荷分析可知(表3),2013年與PC1顯著相關的微生物PLFA生物標記類型的有5種,2014年PC1顯著相關的微生物PLFA生物標記類型增至7種。處理后1年,真菌(20:1w9c)對PC1的貢獻作用降低,細菌(14:0,16:0,cy17:0,i14:0)對PC1的貢獻作用增大。2013年真菌生物標記(20:1 w9c)與PC1極顯著相關,而在2014年與PC1極顯著相關的生物標記物變為革蘭氏陰性細菌(16:1w7c)。不同處理時間,對PC2起顯著作用的微生物PLFA生物標記物類型均有4種,細菌對PC2起極顯著作用,只是標記物類型因處理時間而有所改變(表3)。處理方式對放線菌和革蘭氏陽性細菌影響顯著,處理時間對革蘭氏陰性細菌和總細菌相對豐度影響顯著,處理時間和處理方式對放線菌相對豐度存在顯著的交互作用(表4)。
3.3 不同處理方式對土壤性質的影響
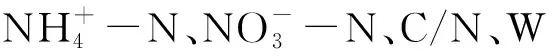
3.4 土壤微生物群落結構與土壤理化性質
對4種處理2個觀測時間的土壤微生物群落結構與土壤理化性質的冗余度分析(RDA)表明,10個土壤理化性質變量在2013年和2014年分別共同解釋了土壤微生物群落結構變化的77.9%和76.5%(圖4)。蒙卡洛檢驗結果顯示,土壤溫濕度比值、土壤濕度是顯著影響2013年微生物群落結構的主要因子;土壤濕度、土壤微生物熵是顯著影響2014年微生物群落結構的主要因子。結合微生物群落結構主成分載荷因子分析可知(表3),不同處理時間,土壤濕度均與主成分分析中PC1顯著相關的微生物PLFA生物標記類型呈現顯著負相關關系,而土壤溫濕度比、MBC/SOC與其呈現顯著正相關關系(圖4)。
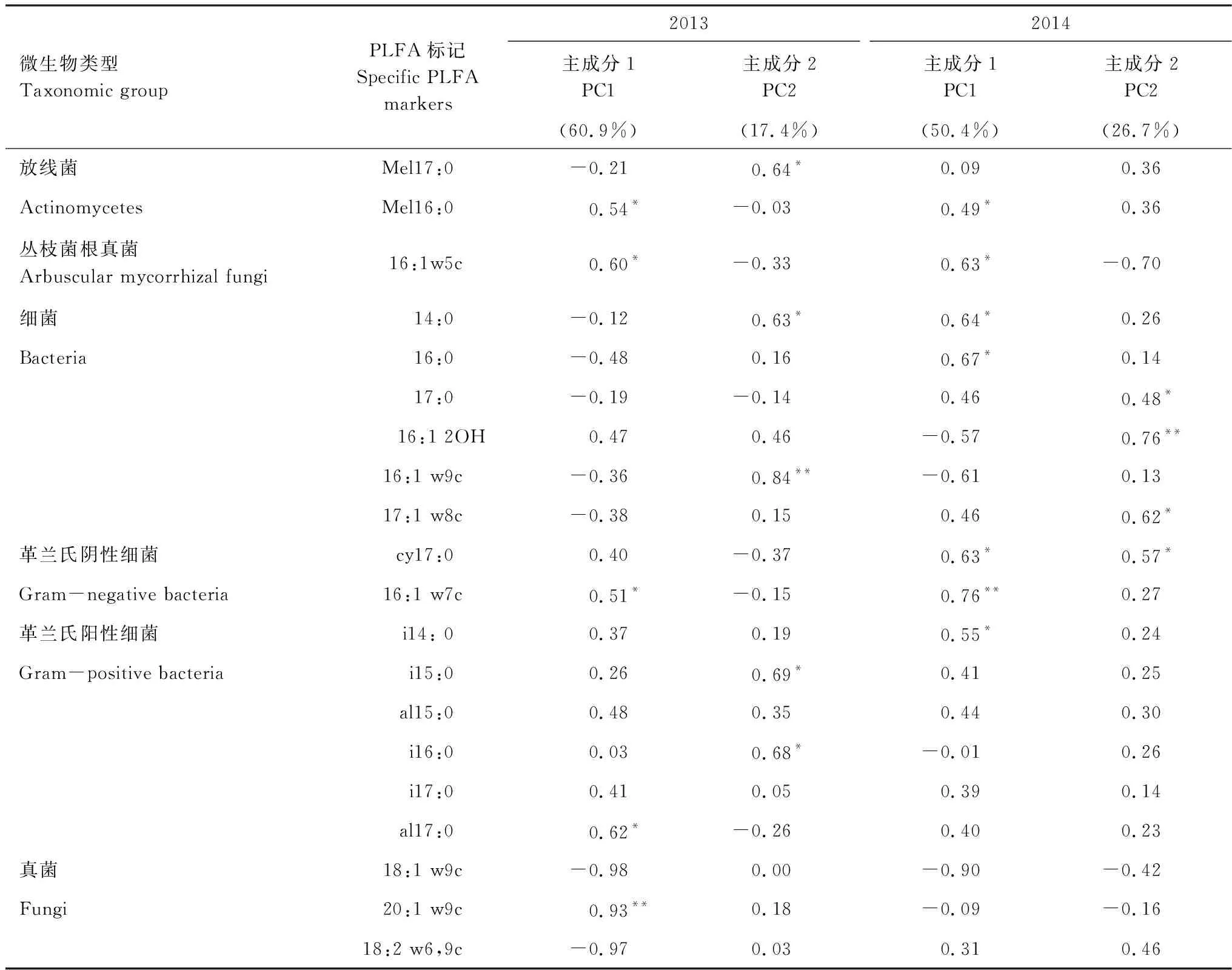
表3 主成分載荷因子
*,**分別表示在0.05和0.01水平下顯著相關
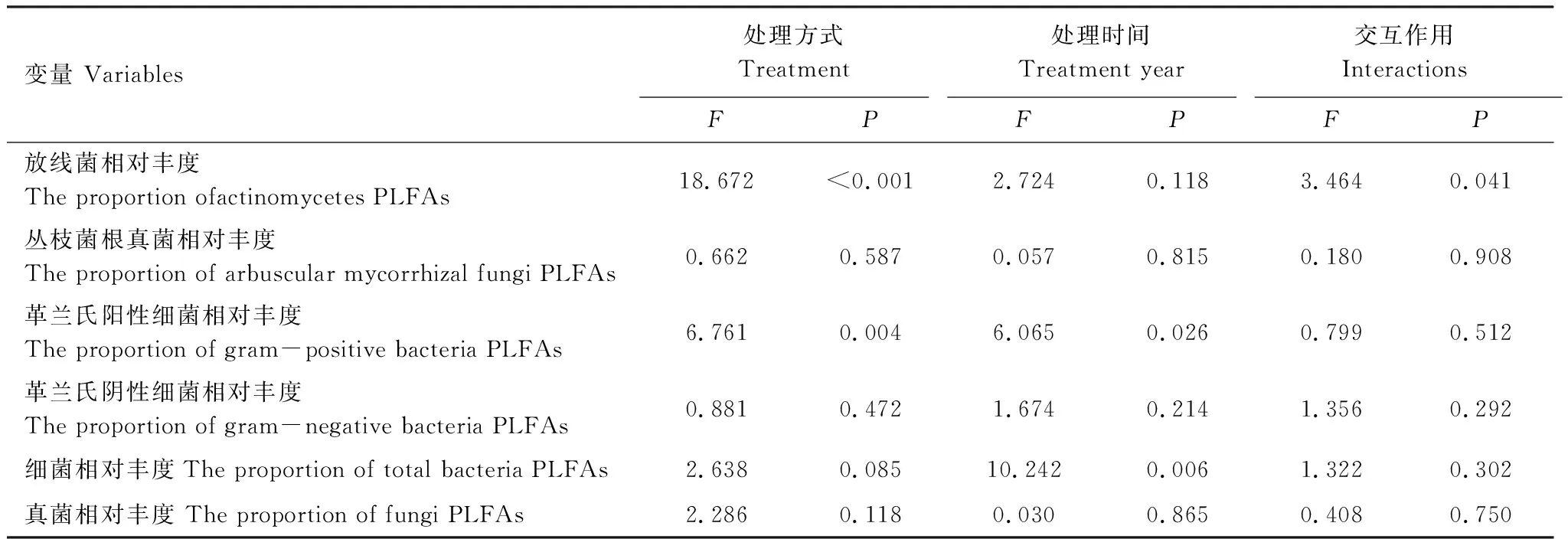
表4 處理方式、處理時間及其交互作用對土壤微生物群落結構的影響
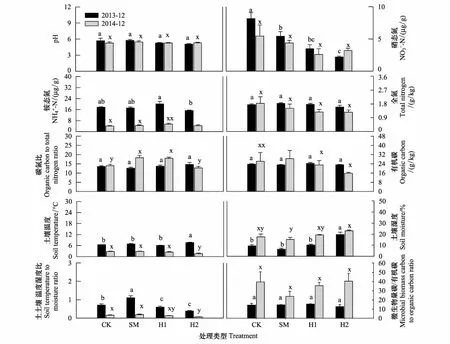
圖3 土壤理化性質對不同處理的響應
4 討論
營林干擾通常通過改變地上底物和地下底物的數量和質量來影響微生物群落,采伐通常會造成針葉林土壤微生物生物量的降低[20- 22]。在本研究中,干擾初期處于不穩定狀態,各處理微生物群落生物量并未呈現出規律性的變化(圖1);但在2014年12月,林內環境逐漸恢復穩定后,除了AMF群落外,SM、H1、H2的其他類群生物量的均呈現出降低的趨勢,從而造成了微生物群落總生物量下降的趨勢。其中,真菌是有機質分解和形成的主要驅動因子[6, 23],一般呈塊狀分布,具有較高的空間異質性[13],而較高的空間異質性使其更易受到采伐等干擾的影響[24],這可能是SM、H1和H2的真菌生物量在處理后1年多呈現出降低趨勢的原因(圖1)。但采伐干擾后初期,死根的分解以及采伐殘余物的輸入可能會引起激發效應,刺激真菌類群,大量以寄生、腐生為營養方式的真菌增加,引起真菌類群的增加[6],采伐后初期SM、H1和H2的真菌生物量的高于CK可能源于上述原因。干擾后真菌類群生物量的變化反映了底物供應對真菌類群的影響過程。多數細菌以真菌的分解產物為主要底物[6],真菌類群的改變也會引起細菌的變化。本研究中,不同處理時間,PC1的極顯著載荷因子由真菌PLFA生物標記物(20:1w9c)轉變為革蘭氏陰性細菌PLFA生物標記物(16:1w7c)(表3),因此,真菌群落結構的變化可能是造成本研究中不同處理時間微生物群落結構變化的主要類群(圖2)。而真菌類群中的AMF與土壤濕度密切相關,高的土壤濕度會抑制AMF的生長[25],這與本研究結果一致,AMF與濕度顯著負相關(圖4),但各處理AMF生物量并未隨著土壤濕度的變化而出現顯著差異(圖1),可能是采伐后其他原因造成的,仍需進一步分析。
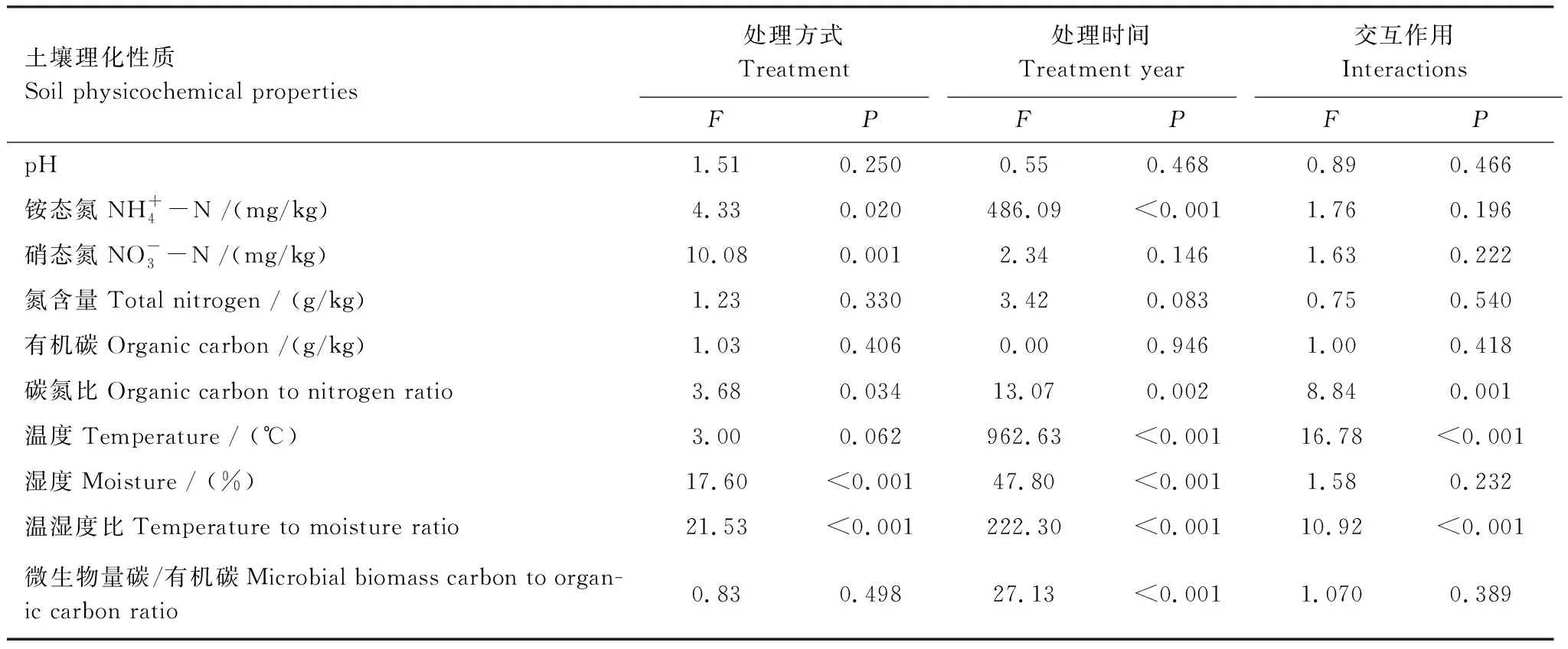
表5 處理方式、處理時間及其交互作用對土壤理化性質的影響
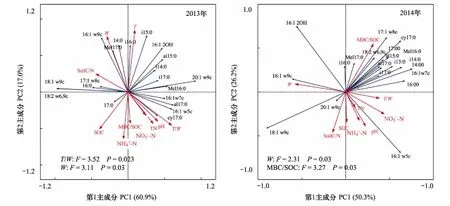
圖4 不同處理時間土壤微生物磷脂脂肪酸與土壤理化性質冗余度分析
相較于真菌,細菌對于環境因子的變化(如土壤溫度、濕度、通氣度等)適應范圍更大[26- 27],同時對于土壤溫濕度變化更為敏感[28]。其中,放線菌能夠通過生產相容物如脯氨酸、甘油等,保持細胞膨脹,抵抗在水分壓力下的質壁分離,同時其絲狀特征能夠有效利用土壤中的水分和空氣[29],因此在土壤溫濕度比值較高的情況下,ACT生物量占優勢,初期SM的ACT生物量高于其他處理(圖1),可能是由于較高的溫濕度比值造成的(圖3)。1年后,H1 和H2 的ACT顯著低于CK(圖1),其溫濕度比低于CK和SM可能是造成ACT變化的主要原因(圖3)。
地上部分和地下部分的群落結構具有強烈的交互作用,植被類型以及組成是影響微生物群落結構主要因子之一[30- 31],SM、H1和H2均改變了地上植被組成以及凋落物的屬性,從而影響微生物群落結構的底物供應,因此處理后1年,這三種措施的微生物群落結構顯著區別于CK(圖2)。而上層植被干擾(H1和H2)和下層植被(SM)屬于兩種不同的干擾方式,盡管灌叢生物量相較于上層植被要小很多,但卻是微生物的重要底物來源[30],因此灌叢的移除極可能通過影響底物組成來改變微生物群落結構的組成(圖2)。相關研究認為由于針葉樹種和闊葉樹種的提供的養分不同,影響共生真菌類群而顯著影響土壤微生物群落組成[32],但那是在改變優勢樹種屬性的前提下[33],本研究中的兩種采伐處理雖然影響了針葉樹種和闊葉樹種的組成,但優勢樹種仍是馬尾松,因此微生物群落結構以及叢枝菌根真菌在兩個觀測期并無顯著差異(圖1,圖2)。不同的微生物種類對土壤有機碳的降解能力不一樣,導致土壤有機碳周轉速率的不同[34],這與本研究的前期研究結果一致。由前期研究可知,與CK相比,H1和H2顯著增加了礦質土壤呼吸,而SM顯著降低了礦質土壤呼吸,微生物群落結構的變化可能是造成土壤呼吸主要組分礦質土壤呼吸變化的重要因素[15]。

5 結論
土壤微生物群落對于森林采伐等干擾措施的響應直接調控著森林土壤碳固定與轉化,本研究通過對采伐處理后不同時期馬尾松林的土壤微生物生物量、群落結構以及相關環境因子進行了分析,結果表明,處理后初期各處理土壤微生物群落生物量以及微生物群落結構并無顯著差異,而在處理后1年多,除灌、采伐1和采伐2 微生物生物量均呈現降低的趨勢,且PCA分析結果顯示,處理后1年多,林下植被剔除以及上層采伐均顯著改變了土壤微生物群落結構。微生物群落結構的變化可能是不同處理土壤異氧呼吸變異的主要原因,但具體機制仍需進一步分析進行確定。同時本研究對不同采伐處理后的相關環境因子進行了探討,土壤溫濕度、土壤微生物熵是造成微生物群落結構變化的關鍵因子,但本研究不同時期結果的差異性表明營林措施的效應會隨時間而改變,因此下一步的研究要注重營林干擾的持續性的作用。
猜你喜歡
小獼猴智力畫刊(2023年4期)2023-04-23 08:49:58
哲學評論(2021年2期)2021-08-22 01:53:34
中華詩詞(2019年7期)2019-11-25 01:43:04
模具制造(2019年3期)2019-06-06 02:10:54
中學生數理化·高一版(2018年1期)2018-02-10 05:20:03
影視與戲劇評論(2016年0期)2016-11-23 05:26:01
七彩語文·寫字與書法(2016年7期)2016-07-28 21:40:22
七彩語文·寫字與書法(2016年6期)2016-07-15 19:36:34
人間(2015年21期)2015-03-11 15:23:21
現代企業(2015年9期)2015-02-28 18:56:50
