基于染色體片段置換系的野生稻粒長QTL——qGL12的精細定位
2018-10-11 02:15:12丁膺賓張莉珍許睿王艷艷鄭曉明張麗芳程云連吳凡楊慶文喬衛華蘭進好
中國農業科學 2018年18期
關鍵詞:水稻
丁膺賓,張莉珍,許睿,王艷艷,鄭曉明,張麗芳,程云連,吳凡,楊慶文,喬衛華,蘭進好
?
基于染色體片段置換系的野生稻粒長QTL——的精細定位
丁膺賓1,2,張莉珍1,2,許睿2,王艷艷2,鄭曉明2,張麗芳2,程云連2,吳凡2,楊慶文2,喬衛華2,蘭進好1
(1青島農業大學農學院,山東青島 266109;2中國農業科學院作物科學研究所,北京 100081)
【目的】利用野生稻染色體片段置換系以及次級群體,精細定位與水稻粒長相關的QTL,發掘野生稻中影響粒長的新基因,為水稻育種提供遺傳材料和基因資源。【方法】利用中國農業科學院作物科學研究所野生稻實驗室已構建的野生稻染色體片段置換系群體,定位到一個與粒長相關的QTL——。在此基礎上,選擇粒長與受體親本9311差異顯著,且攜帶片段的置換系CSSL141與9311回交構建次級分離群體,設計區間內分子標記引物,篩選交換單株,結合粒型表型分析與基因型鑒定,對進行精細定位。通過掃描電子顯微鏡檢測穎殼細胞,觀察細胞的長寬變化,對定位區間內的基因進行基因注釋以及測序分析,預測候選基因。【結果】根據整套置換系多個環境下的表型鑒定,初定位于第12染色體標記RM28621附近;選取攜帶的置換系CSSL141作為精細定位的親本,CSSL141攜帶4個野生稻導入片段,在多年多點的田間試驗中,CSSL141粒長、粒寬、粒重均明顯高于9311;利用構建的CSSL141/9311 F2群體將定位于第12染色體標記RM5479與RM28621之間,影響粒長、粒寬以及粒重,對粒長的貢獻率最高,為44.61%。在定位區間內設計了7個多態性分子標記引物,選擇目標區間基因型雜合的植株種植F3,通過篩選交換單株,結合交換單株的基因型與表型,將定位于RM5479與RM28586之間50 kb區間內,為進一步縮短定位區間,在此區間內設計了4個多態性分子標記引物,選擇交換單株種植下一代,篩選后得到20株交換單株,結合交換單株基因型與籽粒表型,最終將定位到第12染色體15.69 kb區間,該區間內有3個候選基因,其中編碼一種微管蛋白,編碼一種質膜鈣轉運ATP酶,尚未有明確功能,通過測序分析發現,、在編碼區內存在變異;對親本以及后代交換單株的穎殼細胞進行電鏡掃描,發現9311穎殼細胞的長度與寬度均比CSSL141小CSSL141/9311 F4代群體中,目標區間為野生稻基因型的交換單株穎殼細胞的長度與寬度均比目標區間為9311基因型的交換單株大,表明通過調控穎殼細胞的大小影響水稻粒長。【結論】利用野生稻染色體片段置換系,將野生稻粒長QTL——定位于第12染色體15.69 kb區間內,通過調控水稻穎殼細胞的大小影響粒長。該區間內有3個基因。來自野生稻的與與栽培稻等位基因相比,存在自然變異,確定為的候選基因。
野生稻;染色體片段置換系;粒型;QTL定位
0 引言
【研究意義】普通野生稻中含有水稻馴化過程中丟失或者削弱的大量優異基因;粒長是水稻首選的育種目標之一。利用以普通野生稻為供體親本構建的染色體片段置換系進行粒長QTL定位,可以挖掘出野生稻中的優異基因,為水稻育種以及粒長形成的分子機理研究提供新的基因資源。【前人研究進展】水稻是世界上重要的糧食作物之一[1]。中國是稻米消耗大國,65%以上的人口以大米為主食[2]。提高水稻產量對確保中國糧食安全和社會穩定發展有重要意義。水稻產量主要由3個因素構成:單株穗數、每穗粒數和粒重。粒重指標由粒長、粒寬、長寬比和粒厚來綜合評價,這四個性狀與千粒重成正相關[3-4]。因此,籽粒形狀是水稻首選的育種目標之一。與大多數農藝性狀一樣,水稻粒型是受多基因控制的數量性狀[5-6],它不但影響水稻的產量,還影響加工品質與外觀品質。對籽粒形狀相關數量性狀位點(quantitative trait locus,QTL)的鑒定以及新基因的發掘是水稻育種與遺傳研究的重要目標。隨著分子生物學技術以及現代遺傳學的發展,許多研究者通過全面解析粒型相關的遺傳機制和挖掘功能基因/QTL位點,定位了越來越多的粒型QTL,并克隆出一些控制粒型性狀的重要基因。林鴻宣等[7]利用RFLP圖譜對控制秈稻粒型性狀QTL進行定位,在特三矮2號/CB1128群體中定位到5個控制粒長QTL;在外引2號/CB1128群體中定位到5個控制粒長QTL,5個控制粒寬QTL。近年來,很多研究通過構建染色體片段置換系進行水稻優異基因的挖掘定位。周麗慧等[8]通過9311/日本晴CSSL群體定位到10個QTL,與單株產量、單株實粒數、千粒質量、每穗穎花數、結實率、單株總穎花數6個性狀相關。朱文銀等[9]利用染色體片段置換系定位到3個控制水稻粒型的主效QTL,其中有2個粒長相關的QTL,分別為和。被定位在水稻第3染色體RM5551與RM6832之間,遺傳距離為14.8 cM;被定位在RM6832與RM3513之間,其遺傳距離為5.3 cM。隨著第一個控制水稻粒型的基因3[10]被克隆,許多控制粒型的基因如2[11]、2[12]、2[13]、3.1[14]、5[15]、5[16]、7[17]和[18]等相繼被克隆,這些研究為理解調控粒型的機理提供了理論基礎,也對作物的遺傳改良具有重要意義。【本研究切入點】目前,很多研究都是以栽培稻為研究對象,被挖掘出的QTL大多來源于栽培稻,對于野生稻中優異基因的發掘鮮見報道。野生稻異質性強,構建合適的遺傳群體是對其利用的關鍵。構建一套可以永久性利用的染色體片段群體,不僅建立了一個野生稻基因組學研究的平臺,還可以為水稻育種提供新的種質資源。【擬解決的關鍵問題】本研究以海南普通野生稻為供體親本,9311為受體親本,利用中國農業科學院作物科學研究所野生稻實驗室已構建的染色體片段置換系群體,通過多年多點表型鑒定試驗,選擇粒型差異明顯的置換系構建次級群體,設計分子標記結合基因型與表型鑒定進行精細定位,發掘與水稻粒長相關的QTL,尋找野生稻中影響粒長的新基因。
1 材料與方法
1.1 試驗材料
以中國農業科學院作物科學研究所野生稻實驗室構建的一套野生稻染色體片段置換系為研究材料。以來自海南的普通野生稻材料為供體親本,栽培稻品種9311為受體親本,使用119個SSR標記和62個Indel標記,通過多代回交與分子標記輔助選擇,構建出覆蓋水稻染色體的200個染色體片段置換系[19]。齊蘭等[19]利用構建好的置換系群體定位到了37個粒型相關QTL,其中粒長相關QTL——定位于RM28621標記附近,且能在4個環境中檢測到。選擇粒長與受體親本9311差異顯著,且攜帶片段的置換系CSSL141與9311回交,構建次級分離群體,分別于2016年南京、2016年三亞、2017年北京和2017年南京進行種植進行粒長QTL——的精細定位。
1.2 田間種植
2016年5月在南京基地種植200株CSSL141/9311 F2群體作為初定位群體,10月單株收獲籽粒檢測籽粒表型。從初定位結果中挑選出在定位區間內存在基因型差異,而其他區間的基因型為父本或母本的純合基因型的單株于2016年12月在海南試驗基地種植得到400株F3群體,2017年4月收獲F3單株,篩選交換單株縮短定位區間,2017年5月從F3中選擇目標區間雜合基因型的種子在北京、南京試驗基地種植,分別得到2 400株、1 200株F4群體,2017年10月單株收獲F4群體籽粒。田間種植規格為,行距16 cm×26 cm,每行種植10株。
1.3 表型鑒定
水稻成熟后單株收獲種子,自然曬干,籽粒的表型鑒定均用萬深SC-G自動考種及千粒重儀(杭州萬深檢測科技有限公司)進行,測定籽粒的粒長、粒寬以及千粒重。每份材料2次重復,每次測量150—200粒種子。
1.4 群體基因型鑒定
所用的SSR引物來源于美國Cornell大學公布的研究成果,定位區間內引物的設計是根據已經測序完成的日本晴和9311的序列信息,使用primer5軟件進行引物設計,在Gramene網站進行在線比對,驗證其所在染色體位置,確保標記的準確性。利用PCR擴增檢驗在兩親本間的多態性,選擇多態性良好的引物作為精細定位的標記引物。試驗自行設計的定位引物序列見表1。
1.5 粒型性狀QTL定位
根據SSR分析的結果,與親本CSSL141一致的帶型記為2,與9311帶型一致的帶型記為0,同時具有兩親本的雜合帶型記為1,由于某種原因造成帶型缺失或模糊的記為-1。用QTL Ici-Mapping V4.1軟件中的完備區間-加性模型(ICIM-ADD)定位方法,掃描步長為1 cM,以LOD值2.5作為閥值,在全基因組范圍內進行掃描,判斷可能存在的控制粒型性狀的QTL,QTL的命名遵循McCouch等的原則。
1.6 穎殼細胞掃描電鏡觀察
選取9311、CSSL141籽粒,F4群體中定位區間基因型分別為野生稻、9311植株的籽粒,將穎殼分離,剪取中間部分的外穎,固定后將穎殼外穎進行噴金鍍膜,使用掃描電鏡觀察穎殼外表面表皮細胞,檢測穎殼細胞的長度與寬度。
1.7 候選基因預測
根據QTL精細定位結果,利用Rice Genome Browser網站(http://rice.plantbiology.msu.edu/cgi-bin/ gbrowse/rice/#search)查找區間內的所有基因,通過Rice Genome Annotation Project對基因注釋進行分析,篩選出與目標性狀相關的候選基因。在NCBI數據庫(https://www.ncbi.nlm.nih.gov/)中下載候選基因序列,用Primer5設計測序引物對候選基因進行擴增、測序。比較候選基因在9311與CSSL141間的序列差異。
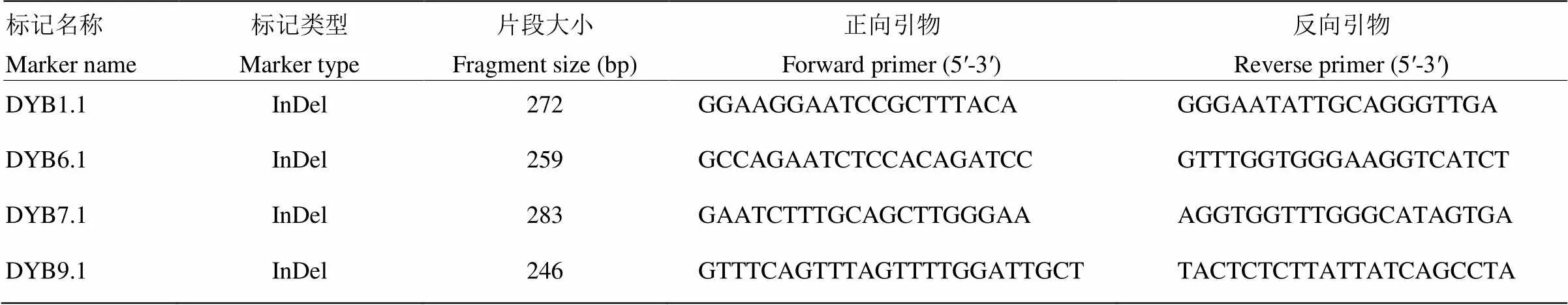
表1 用于精細定位的引物序列
2 結果
2.1 親本CSSL141的選擇以及基因型與表型分析
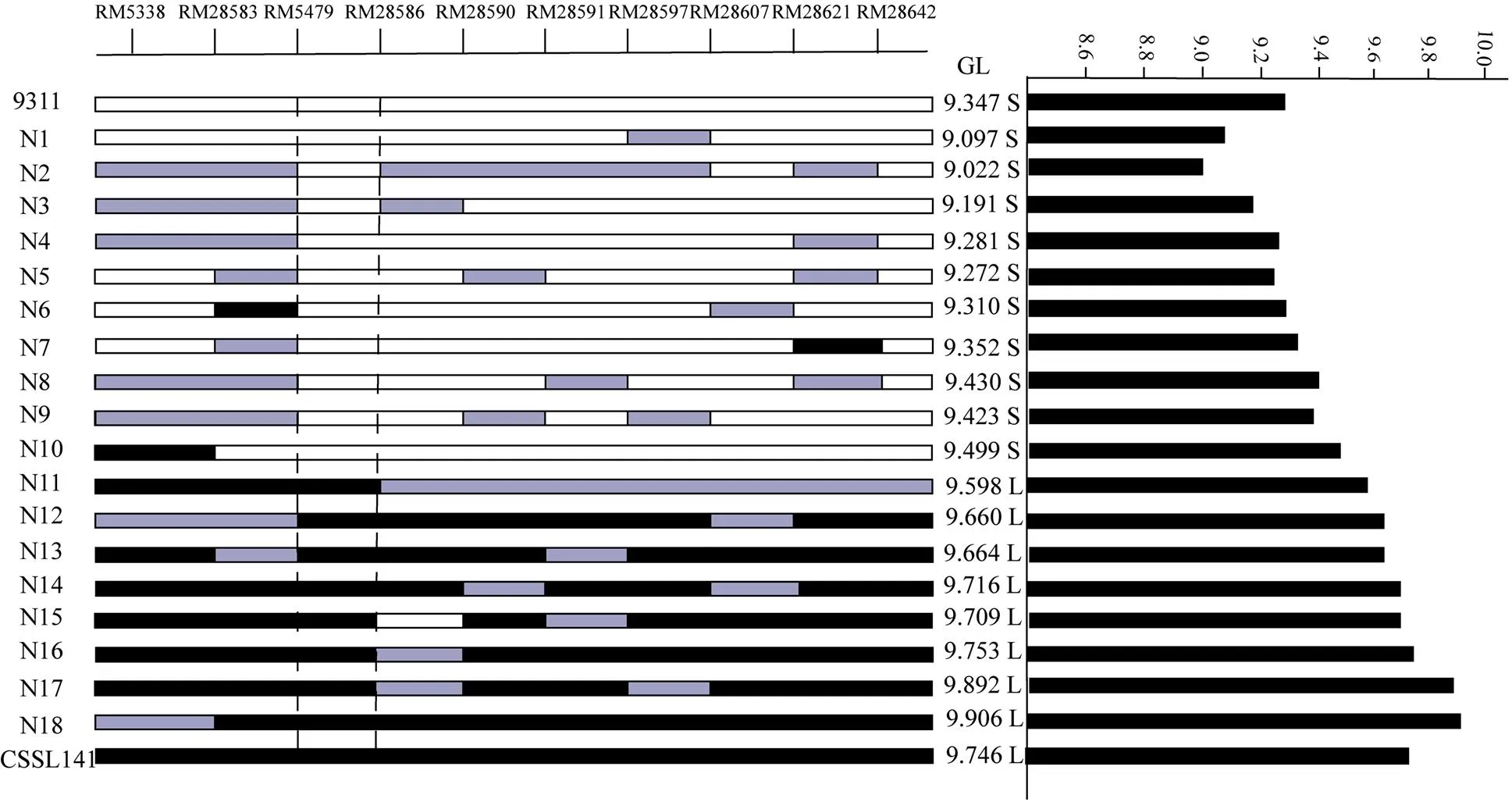
利用染色體片段置換系群體,在多個環境下進行了粒型QTL的定位[19]。在第12染色體RM28621標記附近檢測到一個能夠在多個環境下穩定表達,與粒長相關的QTL,命名為(表2)。選擇攜帶片段且粒型與9311差異顯著的置換系CSSL141作為親本,進行下一步的精細定位。CSSL141染色體基因型如圖1所示,攜帶4個來自野生稻的置換片段,1個片段位于第2染色體,3個片段位于第12染色體,其中第12染色體上的一個置換片段位于RM28621標記附近(圖1)。
根據整套置換系材料在2014—2016年3年不同環境中的調查結果,發現CSSL141和親本9311在粒型上有明顯差異(圖2-a和圖2-b)。其中,9311在北京、南京、三亞粒長均值分別為9.61、9.77和9.35 mm,CSSL141粒長均值分別為10.10、10.19和9.90 mm;9311千粒重均值分別為31.22、32.60和32.22 g,CSSL141千粒重均值分別為33.83、35.04和35.08 g;粒長、千粒重差異均達到極顯著水平(<0.001)。 9311粒寬均值在北京、南京、三亞分別為2.79、2.79和2.83 mm,CSSL141粒寬均值分別為2.82、2.86和2.93 mm(圖2-c)。此外,CSSL141與9311在株高、穗長、有效穗數等農藝性狀方面差異不顯著。CSSL141中置換片段較少,籽粒表型與9311差異較大而其他農藝性狀與9311沒有顯著差異,而且其置換片段中含有實驗室已經定位到的QTL——,因此選為親本進行的精細定位。
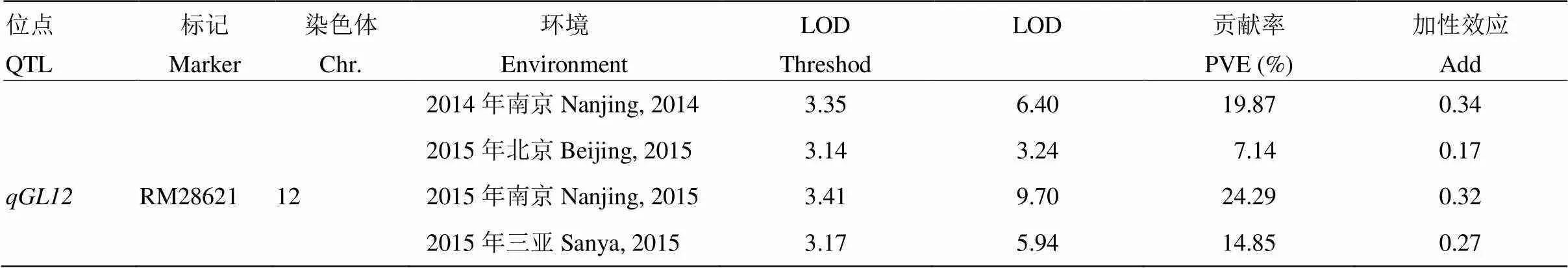
表2 不同環境下qGL12的檢測
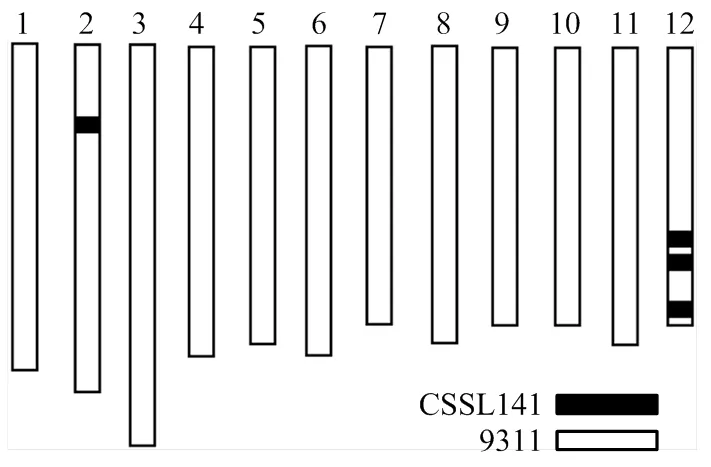
圖1 CSSL141置換片段分布
2.2 F2群體定位結果
CSSL141/9311 F2群體種植于南京試驗基地。粒長、粒寬以及千粒重3個粒型性狀在F2群體的分布均呈連續正態分布(圖3)。F2群體粒長介于9.58—10.80 mm,主要集中在9.81—10.27 mm;粒寬介于2.57—2.84 mm,主要集中在2.68—2.72 mm;千粒重介于30.50—36.63 g,主要集中在32.76—35.12 g。說明這三個性狀是由多基因調控,屬于數量遺傳性狀,適合QTL分析。
根據CSSL141染色體基因組中野生稻置換片段的分布,使用置換片段區間或周圍的分子標記設引物RM5338、RM5479、RM28621、RM28642、Indel12-12作為初定位標記引物。利用QTL IciMapping分析軟件將定位于第12染色體標記RM5479—RM28621,影響粒長、粒寬以及粒重。對粒長的貢獻率最高,為44.61%(表3)。
2.3 qGL12精細定位
利用F2群體將定位于第12染色體標記RM5479與RM28621之間。F2群體中選擇兩標記基因型雜合的植株進行種植得到F3。在定位區間RM5479—RM28621兩邊及中間增加7個多態性分子標記引物,分別為RM28583、RM28586、RM28590、RM28591、RM28597、RM28607和RM28642。對400株海南F3單株進行基因型分析,篩選出18株交換單株。結合交換單株粒長表型與基因型,將定位于RM5479和RM28586之間,區間長度約為50 kb(圖4和圖5)。
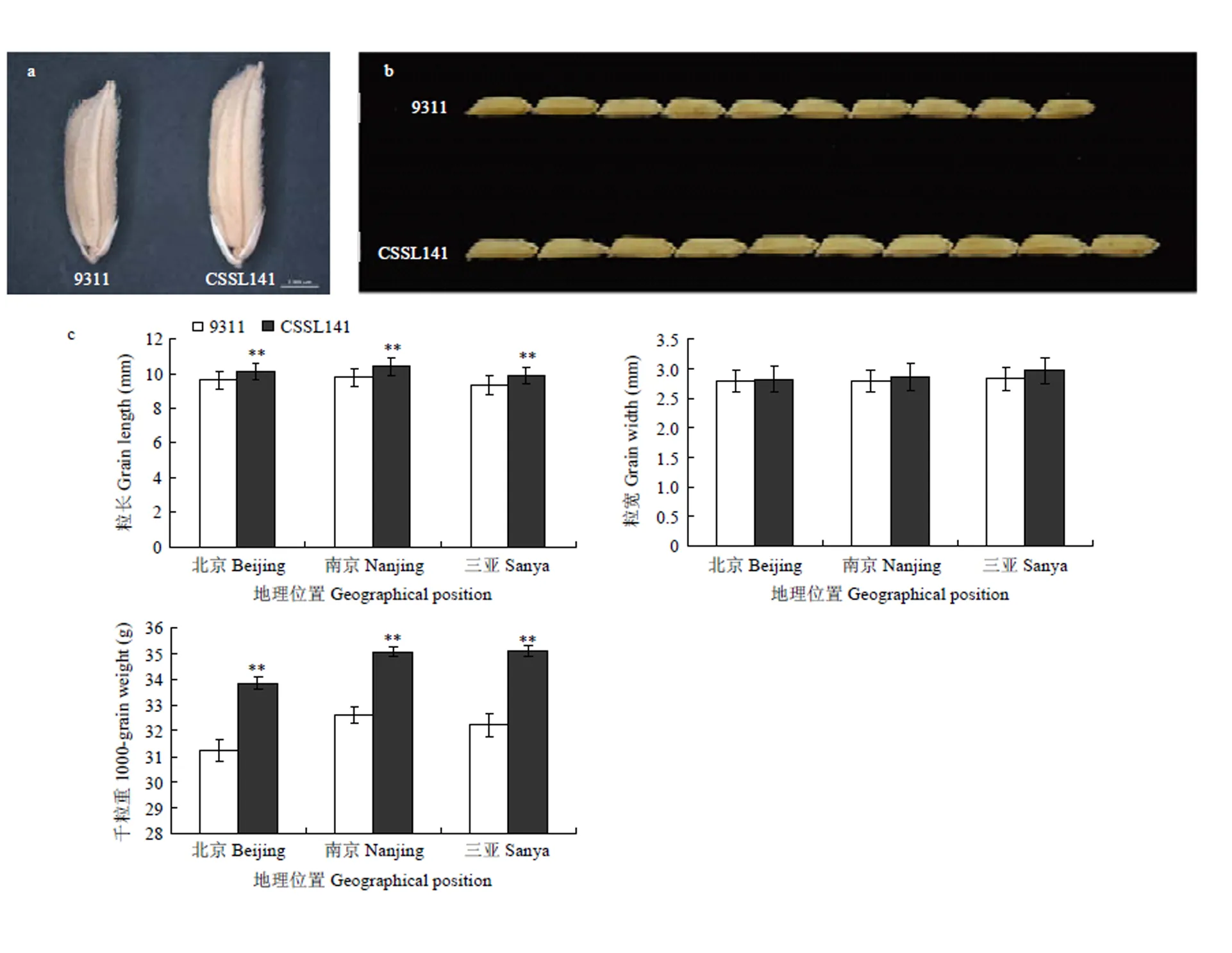
a:單粒種子;b:粒長比較;c:不同環境下的粒型性狀比較。**差異極顯著(<0.001)
a: The seeds of 9311 and CSSL141; b: 10 Comparison of grain length; c:Comparison of grain traits under multiple environments. **significantly different at<0.001
圖2 9311與CSSL141籽粒性狀比較
Fig. 2 Comparison of kernel traits between 9311and CSSL141
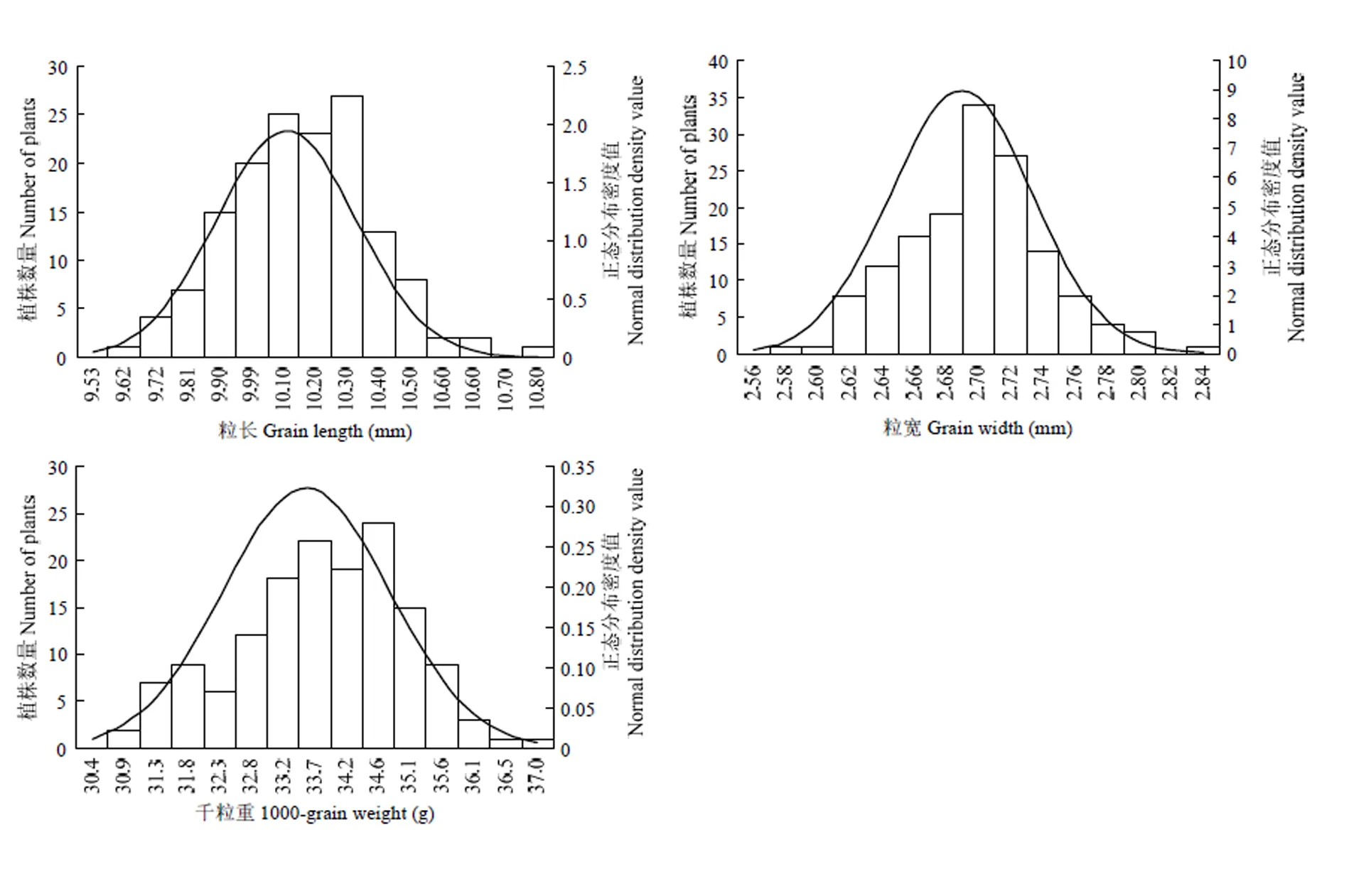
圖3 F2群體粒長、粒寬、千粒重頻次分布圖
Fig. 3 Frequency distribution diagram of grain length, grain width and 1000 grain weight in CSSL141/9311 F2population
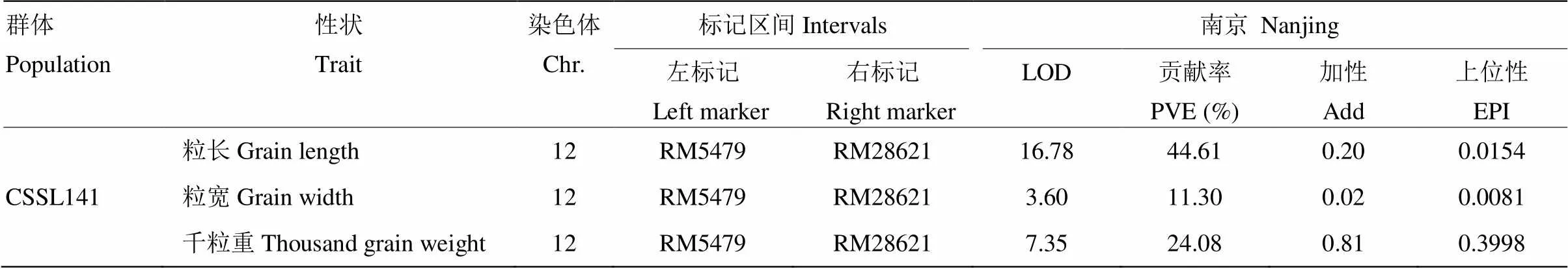
表3 CSSL141/9311 F2群體粒型QTL初定位結果
為了進一步精細定位,將F3群體中目標區間基因型雜合的單株種子分別在北京順義、南京進行種植得到F4。在NCBI數據庫(https://www.ncbi. nlm.nih.gov/)中找到RM5479與RM28586區間片段的序列,利用Primer5軟件設計分子標記引物,設計好的引物在兩親本間進行多態性檢測,最后得到4個多態性良好的分子標記引物DYB1.1、DYB6.1、DYB7.1和DYB9.1(表1)。對2 400株種植于北京順義的F4單株以及1 200株種植于南京的F4單株進行定位分析,南京和北京的群體都在RM5479和RM28586之間發現粒長相關的QTL。通過篩選交換單株,得到20株交換單株,結合基因型與表型的結果,根據交換單株信息,將的左邊界定于DYB9.1標記處,右邊界定于RM28586,區間長度約15.69 kb(圖4和圖6)。
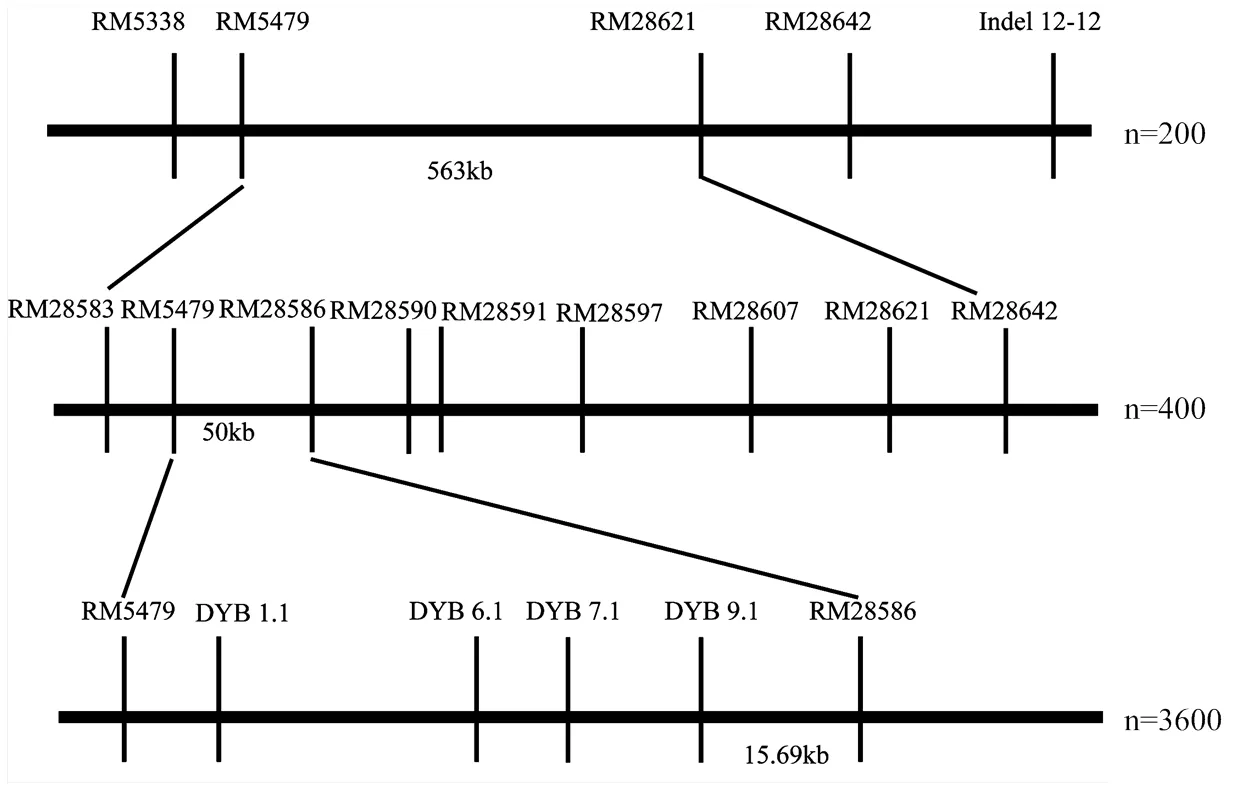
圖4 qGL12的精細定位
2.4 穎殼細胞學觀察
取同一時期、同一環境下9311與CSSL141的籽粒,以及F4群體中2個交換單株的籽粒,2個交換單株目標區間染色體片段分別為野生稻基因型以及9311基因型。掃描電鏡觀察穎殼的外表皮細胞。結果顯示,在400×的視距下,親本CSSL141的平均細胞長度為40.82 μm(圖7-a);9311平均細胞長度為34.48 μm(圖7-b);F4交換單株籽粒中標記區間為野生稻基因型的籽粒平均細胞長度為46.51 μm(圖7-c);標記區間為9311基因型的交換單株籽粒平均細胞長度為35.71 μm(圖7-d)。因此推斷通過調控水稻穎殼細胞大小控制籽粒形狀。
2.5 候選基因預測
最終定位于第12染色體15.69 kb的區間,兩側標記為DYB9.1和RM28586。根據Rice Genome Browser網站(http://rice.plantbiology.msu.edu/cgi-bin/ gbrowse/rice/#search)序列分析,該區間存在3個候選基因,分別為、和。
S:短粒;L:長粒;白色區域:9311基因型;黑色區域:野生稻基因型;灰色區域:雜合基因型。下同
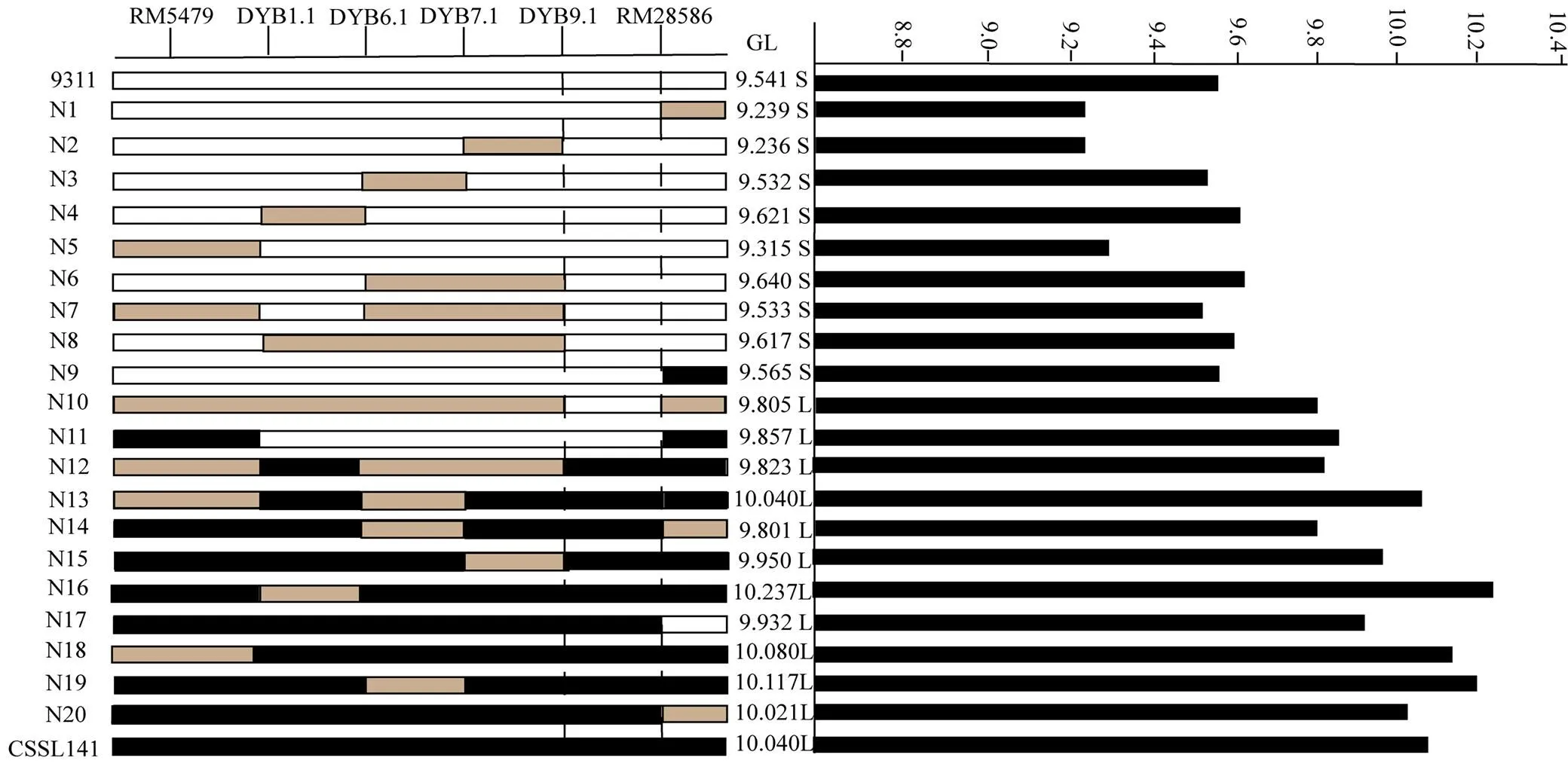
圖6 RM5479與RM28586區間內交換單株基因型與表型
Rice Genome Annotation Project網站基因注釋為:編碼一種微管蛋白,編碼一種質膜鈣轉運ATP酶,尚未有明確功能(表4)。將9311與CSSL141中3個候選基因的測序結果進行對比,在兩親本編碼區中有8個SNP,其中5個導致氨基酸發生改變;第403個堿基中T/A堿基發生替換導致半胱氨酸替換成絲氨酸;第427個堿基中G/T堿基發生替換導致天冬氨酸替換成酪氨酸;第460個堿基中G/A堿基發生替換導致纈氨酸替換成異亮氨酸;第1 450個堿基中G/A堿基發生替換導致丙氨酸替換成蘇氨酸;第1 672個堿基中T/C堿基發生替換導致半胱氨酸替換成精氨酸。在兩親本編碼區中有17個SNP,其中有2個導致氨基酸發生改變;第433個堿基中T/C堿基發生替換導致半胱氨酸替換成精氨酸;第2 754個堿基中T/A堿基發生替換導致天冬氨酸替換成谷氨酸。

a:親本CSSL141;b:親本9311;c:標記區間為野生稻基因型的交換單株;d:標記區間為9311基因型的交換單株

表4 qGL12定位區間內候選基因
3 討論
3.1 染色體片段置換系在水稻QTL定位中的應用
傳統的QTL定位一般采用的是初級定位群體,例如RIL等,受遺傳背景的影響,QTL定位的區間都很大,通常是10 cM以上,難以確定 QTL的作用效應是單個主效QTL的作用還是多個微效QTL的作用。對于精細定位,最關鍵的環節就是構建合適的定位群體,篩選與目標基因緊密連鎖的分子標記。理想的定位群體是除了目標基因所在座位的局部區間外,其他部分的遺傳背景是相同的[20]。染色體片段置換系僅在置換片段上發生分離,其余遺傳背景與受體親本完全一致,因此,消除了遺傳背景的干擾,尤其是能夠檢測出遺傳效應較小的QTL,是QTL定位的理想工具[21]。
本研究中,粒型QTL的定位是建立在染色體片段置換系多年多點田間試驗的基礎上,利用構建的置換系群體定位到了37個粒型相關QTL,其中粒長相關的QTL在2年3點多環境試驗中在4個環境中均被檢測到,說明是可以在不同環境下穩定遺傳的QTL。從CSSL群體中挑選粒型差異與9311較大而且攜帶置換片段的CSSL141與9311進行回交,通過F2、F3以及F4群體進行精細定位。最終將定位于第12染色體15.69 kb的區間,影響粒長、粒寬以及粒重,對粒長的貢獻率最高。利用野生稻染色體片段置換系進行精細定位,能夠充分消除遺傳背景的影響,發掘野生稻中的優異基因,將成為一種主流方法。
3.2 qGL12通過調控穎殼細胞大小影響粒長
稻米的形狀和大小受到穎殼形狀和大小的嚴格控制,這表明穎殼的形態影響水稻的外觀品質和產量,而穎殼的形態則是由細胞的大小、數目以及排列方式決定的。基因影響水稻粒型的方式可能有兩種,一種為影響穎殼細胞的數量。例如[22]通過改變細胞分裂模式來控制籽粒形狀,使縱向細胞分裂增加橫向細胞分裂減少形成細長的籽粒。一種為影響穎殼細胞的大小。如[23]通過調控穎殼細胞的長度與寬度影響水稻粒型;[24]和[25]主要通過影響穎殼細胞的長度影響籽粒形狀。本研究利用掃描電鏡對水稻9311、CSSL141以及2個F4交換單株進行水稻穎殼細胞觀察,發現9311穎殼細胞的長度與寬度均比CSSL141小,2個交換單株中,目標區間為野生稻基因型的交換單株穎殼細胞的長度與寬度均比目標區間為9311基因型的交換單株大,這表明通過調控細胞的長度與寬度影響水稻粒型。
3.3 qGL12應用前景
區間內含有3個基因,3個基因均未見報導與水稻粒長有關,測序結果表明2個候選基因在編碼區均有非同義突變。下一步的研究將利用轉基因等方法對2個候選基因進行功能驗證,確定野生稻中控制粒長的基因。置換系CSSL141,或者CSSL141/9311 F4群體中攜帶的株系,可以直接作為水稻品種改良的育種材料,DYB9.1與RM28586可以作為分子標記用于輔助選擇。本研究將為水稻粒長形成的分子機理研究提供新的基因資源,同時,為水稻粒長的改良提供了新的育種材料。
4 結論
被定位于第12染色體15.69 kb的區間,兩側標記分別為DYB9.1和RM28586。區間內與2個基因在編碼區均有非同義突變,被確定為的候選基因,并且通過影響水稻籽粒穎殼細胞的大小影響粒長。
[1] 景春艷, 張富民, 葛頌. 水稻的起源與馴化——來自基因組學的證據. 科技導報, 2015, 33(16): 27-32.
Jing C Y, Zhang F M, Ge S. Genomic evidence of the origin and domestication of Asian cultivated rice (L.)., 2015, 33(16): 27-32. (in Chinese)
[2] Khush G S. What it will take to feed 5.0 billion rice consumers in 2030., 2005, 59: 1-6.
[3] 涂坦, 付洪. 水稻主要粒型基因及其遺傳調控的研究進展. 山地農業生物學報, 2016, 35(5): 58-65.
Tu T, Fu H. Research progress in major genes controlling grain shape of rice and their genetic regulation., 2016, 35(5): 58-65. (in Chinese)
[4] 梁云濤, 潘英華, 徐志健. 利用野栽分離群體定位水稻粒型相關QTL. 西南農業學報, 2017, 30(10): 2161-2167.
Liang Y T, Pan Y H, Xu Z J. QTLs mapping of grain shape of rice by using offspring derived from cross ofGriff. and cultivated rice., 2017, 30(10): 2161-2167. (in Chinese)
[5] 邢永忠, 談移芳, 徐才國, 華金平, 孫新立. 利用水稻重組自交系群體定位谷粒外觀性狀的數量性狀基因. 植物學報, 2001(8): 840-845.
Xing Y Z, Tan Y F, Xu C G, Hua J P, Sun X L. Mapping quantitative trait loci for grain appearance traits of rice using a recombinant inbred line population., 2001(8): 840-845. (in Chinese)
[6] 余守武, 樊葉楊, 楊長登, 李西明. 水稻第1染色體短臂粒長和粒寬QTL的精細定位. 中國水稻科學, 2008(5): 465-471.
Yu S W, Fan Y Y, Yang C D, Li X M. Fine mapping of quantitative trait loci for grain length and grain width on the short arm of rice chromosome1., 2008(5): 465-471. (in Chinese)
[7] 林鴻宣, 閔紹楷, 熊振民, 錢惠榮, 莊杰云, 陸軍, 鄭康樂, 黃寧. 應用RFLP圖譜定位分析秈稻粒形數量性狀基因座位. 中國農業科學, 1995(4): 1-7.
Lin H X, Min S K, Xiong Z M, Qian H R, Zhuang J Y, Lu J, Zheng K L, Huang N. RFLP mapping of QTLs for grain shape traits in indica rice., 1995(4): 1-7. (in Chinese)
[8] 周麗慧, 張亞東, 朱鎮, 陳濤, 趙慶勇, 姚姝, 趙凌, 趙春芳, 于新, 王才林. 利用染色體片段置換系群體檢測水稻產量相關性狀QTL. 江蘇農業學報, 2015, 31(1): 1-9.
Zhou L H, Zhang Y D, Zhu Z,CHEN T, ZHAO Q Y, YAO S, ZHAO L, ZHAO C F, YU X, WANG C L. Quantitative trait locus detection for rice yield-related traits using chromosome segment substitution lines., 2015, 31(1):1-9. (in Chinese)
[9] 朱文銀, 楊德衛, 林靜, 趙凌, 張亞東, 朱鎮, 陳濤, 王才林. 利用染色體片段置換系定位水稻粒型QTL. 江蘇農業學報, 2008, 3: 226-231.
Zhu W Y, Yang D W, Lin J, Zhao L, Zhang Y D, Zhu Z, Chen T, Wang C L. Substitution mapping of QTLs for grain shape using chromosome segment substitution lines in rice(L.)., 2008, 3: 226-231. (in Chinese)
[10] Fan C C, Xing, Y Z, Mao H L, Lu T T, Han B, Xu C G, Li X H, Zhang Q F., a major QTL for grain length and weight and minor QTL for grain width and thickness in rice, encodes a putative transmembrane protein., 2006, 112(6): 1164-1171.
[11] Hu J, Wang Y X, Fang Y X, Zeng L J, Xu J, Yu H P. A rare allele ofenhances grain size and grain yield in rice., 2015, 10(8): 1455-1465.
[12] Song X J, Huang W, Shi M, Zhu M Z, Lin H X. A QTL for rice grain width and weight encodes a novel RING-type E3 ubiquitin ligase., 2007, 39(5): 623- 630.
[13] Che R H, Tong H M, Shi B H, Liu Y Q, Fang S R, Liu D P. Control of grain size and rice yield by-mediated brassinosteroid responses., 2015, 2(1): 15195.
[14] Qi P, Lin Y S, Song X J, Shen J B, Huang W, Shan J X, Zhu M Z, Jiang L W, Gao J P, Lin H X. The novel quantitative trait locuscontrols rice grain size and yield by regulating Cyclin- T1; 3., 2012, 22(12): 1666-1680.
[15] Weng J F, Gu S H, Wan X Y, Gao H, Guo T, Su N, Lei C L, Zhang X, Cheng Z J, Guo X P, Wang J L, Jiang L, Zhai H Q, Wan J M. Isolation and initial characterization of, a major QTL associated with rice grain width and weight., 2008, 18(12): 1199-1209.
[16] Li Y B, Fan C C, Xing Y Z, Jiang Y H, Luo L J, Sun L, Shao D, Xu C J, Li X H, Xiao J H, He Y Q, Zhang Q F. Natural variation inplays an important role in regulating grain size and yield in rice., 2011, 43(12): 1266-1269.
[17] Wang Y X, Xiong G S, Hu J, Jiang L, Yu H, Xu J, Fang Y X, Zeng L J, Xu E B, Xu J, Ye W J, Meng X B, Liu R F, Chen H Q, Jing Y H, Wang Y H, Zhu X D, Li J Y, Qian Q. Copy number variation at thelocus contributes to grain size diversity in rice., 2015, 47(8): 944-948.
[18] Wang S K, Wu K, Yuan Q B,LIU X Y, LIU Z B, LIN X Y, ZENG R Z, ZHU H T, DONG G J, QIAN Q, ZHANG G Q, FU X D. Control of grain size, shape and quality by, 2012, 44(8): 950-954.
[19] Qi L, Sun Y, Li J, SU L, ZHENG X M, WANG X N, LI K M, YANG Q W, QIAO W H. Identify QTLs for grain size and weight in common wild rice using chromosome segment substitution lines across six environments., 2017, 472: 482.
[20] 朱文銀, 王才林. 作物染色體片段置換系研究進展. 江蘇農業學報, 2008, 24(6): 963-968.
Zhu W Y, Wang C L. A review on chromosomal segment substitution lines in crops., 2008, 24(6): 963-968. (in Chinese)
[21] Qiao W H, Qi L, Cheng Z J, SU L, LI J, SUN Y, REN J F, ZHENG X M, YANG Q W. Development and characterization of chromosome segment substitution lines derived fromin the genetic background ofspp.cultivar 9311., 2016, 17: 580.
[22] Wang S K, Li S, Liu Q, Wu K, Zhang J Q, WANG S S, WANG Y, CHEN X B, ZHANG Y, GAO C X, WANG F, HUANG H X, FU X D. Theregulatory module determines grain shape and simultaneously improves rice yield and grain quality., 2015, 23(3): 12.
[23] Abe Y, MieDa K, Ando T, KONO I, YANO M, KITANO H, IWASAKI Y. The SMALL AND ROUND SEED1 (SRS1/DEP2) gene is involved in the regulation of seed size in rice., 2009, 85(5): 327-339.
[24] Segami S, Kono I, Ando T, YANO M, KITANO H, MIURA K, IWASAKI Y.gene encodes alphatubulin regulating seed cell elongation in rice., 2012, 5(1): 1-10.
[25] Heang D, Sassa H. Antagonistic actions of HLH/bHLH proteins are involved in grain length and weight in rice., 2012, 7(2): e31325.
(責任編輯 李莉)
Fine Mapping of Grain Length Associated QTL,(L.) Using a Chromosome Segment Substitution Line
Ding Yingbin1,2, Zhang Lizhen1,2, Xu Rui2, Wang Yanyan2, Zheng Xiaoming2, Zhang Lifang2, Cheng Yunlian2, Wu Fan2, Yang Qingwen2, Qiao Weihua2, Lan Jinhao1
(1College of Agriculture of Qingdao Agricultural University, Qingdao 266109, Shandong;2Institute of Crop Sciences, Chinese Academy of Agricultural Sciences, Beijing 100081)
【Objective】Fine mapping of a grain length QTL from wild rice using a chromosome segment substitution line and secondary population, exploring new genes affecting grain length and providing genetic materials and gene resources for rice breeding.【Method】Our laboratory had preliminary mapped a grain length related QTL,using the chromosome segment substitution line (CSSL) population. On this basis, we choose one CSSL, CSSL141 which harborssubstitution segment and has significant difference of grain length compared with receptor parent 9311. CSSL141 was backcrossed with 9311. The secondary separation population was constructed for fine mapping of. Detection of length of glumes cells was performed by scanning electron microscope.【Result】CSSL141 has 4 introgressive segments from wild rice, its grain length, grain width and grain weight was significantly higher than 9311 under multi-conditions. Using CSSL141/9311 F2population,was localized to the interval between RM5479 and RM28621 on chromosome 12.affected grain length, grain width and grain weight, and the phenotypic variation explained of grain length was 44.61%. Seven polymorphic molecular marker primers in the location interval were designed, F3plants which had heterozygous genotype in the target interval were investigated.was mapped to a 50 kb region between RM5479 and RM28586. Four polymorphic molecular marker primers were designed in this interval, individuals which harbor heterozygous genotype in this interval were selected for next generation F4. Eventuallywas narrowed to a 15.69 kb region between DYB9.1 and RM28586 on chromosome 12. There are 3 genes in the interval. Two candidate genes,and, which encode a tubulin protein and a calcium-transporting ATPase respectively, with variations in their coding regions. The results of electron microscopes scanning of glumes cell showed that the length and width of 9311’s glumes cell was smaller than CSSL141, indicated thatregulate grain size of rice by controlling the size of glumes cells.【Conclusion】The wild rice grain length associated QTLwas fine mapped to a 15.69 kb region on chromosome12.control grain length via regulate the size of glumes cells. Two candidate genes,and, were found and would be used for further research.
rice; chromosome segment substitution line; grain size; QTL mapping
10.3864/j.issn.0578-1752.2018.18.001
2018-04-11;
2018-05-26
國家自然科學基金(31471471)、作物生物學國家重點實驗室開放課題(2016KF08)、中國農業科學院創新工程、國家重點研發計劃(2016YFD0100101)
丁膺賓,E-mail:913299099@qq.com。通信作者楊慶文,E-mail:yangqingwen@caas.cn。通信作者喬衛華,Tel:010-62186687;E-mail:qiaoweihua@caas.cn。通信作者蘭進好,E-mail:jinhao2005@163.com
猜你喜歡
幼兒100(2023年39期)2023-10-23 11:36:32
青少年科技博覽(中學版)(2022年6期)2022-12-27 19:44:27
中國土壤與肥料(2021年5期)2021-12-12 02:02:11
今日農業(2021年21期)2021-11-26 05:07:00
軍事文摘(2021年22期)2021-11-26 00:43:51
今日農業(2021年14期)2021-10-14 08:35:40
金橋(2021年7期)2021-07-22 01:55:38
今日農業(2020年20期)2020-11-26 06:09:10
文苑(2020年6期)2020-06-22 08:41:52
文苑(2019年22期)2019-12-07 05:29:00
