環境脅迫植物富集γ-氨基丁酸的研究進展
2018-10-16 09:53:50丁俊胄賈才華趙思明張賓佳熊善柏林親錄
食品工業科技 2018年18期
王 斌,丁俊胄,2,賈才華,*,趙思明,*,牛 猛,張賓佳,熊善柏,林親錄
(1.華中農業大學食品科學技術學院,湖北武漢 430070;2.伊利諾伊大學香檳分校食品與人類營養學系,美國伊利諾伊州厄巴納市 61801;3.中南林業科技大學食品科學與工程學院,湖南長沙 410004)
γ-氨基丁酸(Gama-aminobutyric acid,GABA)是一種四碳非蛋白組成氨基酸,在神經系統中是一種重要的神經活性抑制劑,具有諸多功效,例如降低血壓、治療失眠、緩解焦慮、增強免疫力以及增加記憶力等作用[1]。我國衛生部2009年批準γ-氨基丁酸為新資源食品,建議成人每天攝入500 mg以內。隨著人們年齡的增長,人體通過自身代謝生成GABA的能力降低,加之日常飲食中GABA含量攝入量不夠,人體GABA含量較低。
GABA廣泛分布于植物中,但含量普遍較低,當植物受到環境(低氧、低溫、熱、鹽滲透、機械損傷)刺激時,會大量生成GABA。在食品生產中,可通過改變外界環境刺激植物組織,促進GABA的合成,制得高GABA食品,如茶葉、發芽糙米、豆芽等。
高GABA食品能夠有效增加人體GABA含量,對于保持人體健康具有重要意義。本文將從GABA的代謝機制、環境脅迫對GABA積累的影響以及幾種脅迫方式在高GABA食品研發中的應用做一個綜述。
1 γ-氨基丁酸代謝途徑
高等植物代謝積累GABA有兩種途徑。一種是以谷氨酸(L-Glutamine,Glu)作為底物,經谷氨酸脫羧酶(Glutamate decarboxylase,GAD,EC 4.1.1.15)催化生成GABA,該途徑被稱為GABA旁路或GABA支路[2]。另一種則是以多胺(polyamines)為底物,經過二胺氧化酶(DAO)、多胺氧化酶(PAO)催化,可將多胺(腐胺、亞精胺)轉化為GABA,該途徑被稱為多胺降解[3-4]。
1.1 GABA支路
GABA支路廣泛分布于真核生物和原核生物中,是GABA重要的代謝途徑。如圖1所示[1],在細胞質中,谷氨酸首先被GAD催化不可逆生成GABA,隨后GABA由GABA轉氨酶(GABA transaminase,GABA-T)催化轉化為成琥珀酸半醛半醛酸(Succinic acid,SSA),琥珀酸半醛進入線粒體,由琥珀酸半醛脫氫酶(Succinate-Semialdehyde Dehydrogenase,SSADH)氧化生成琥珀酸(Succinic acid,SSC),隨后琥珀酸進入三羧酸循環[2]。
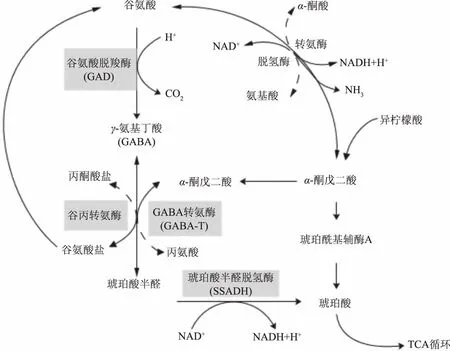
圖1 GABA支路代謝路徑圖Fig.1 GABA shunt and its relationship to other metabolic pathways
1.2 多胺降解
多胺降解也是生成GABA的途徑之一。相較于GABA支路,多胺降解產生的GABA含量較少,在以往的研究中,大部分都是以GAD活性作為關鍵因素來探究GABA在植物體內代謝情況。但最近的研究發現,在某些植物中多胺的含量與GABA含量有密切聯系,通過多胺降解產生的GABA占有較大比例。Yang等[5]用氨基胍對大豆進行處理發現,氨基胍能顯著增加多胺含量,但GABA含量下降了30%。另外,Wang等[6]用外源GABA處理甜瓜根,發現處理后甜瓜根的多胺含量顯著增加,說明多胺代謝與GABA代謝有密切聯系。
多胺降解產生GABA路徑如圖2所示,谷氨酸(L-Glutamic acid,L-Glu)經過轉氨酶生成鳥氨酸(Ornithine,Orn)和精氨酸(Arginine,Arg),鳥氨酸和精氨酸經過鳥氨酸氧化酶(Ornithine decarboxylase,ODC)和精氨酸氧化酶(Arginine decarboxylase,ADC)氧化生成腐胺(Putrescine,Put),隨后腐胺經亞精胺合成酶催化轉化成亞精胺(Spermidine,Spd),亞精胺進一步轉化為精胺(Spermine,Spm)。腐胺和亞精胺分別經二胺氧化酶(Diamine oxidase,DAO,EC 1.4.3.6)和多胺氧化酶(Polyamine oxidase,PAO,EC 1.5.3.3)氧化生成4-氨基丁醛,最后由氨基丁醛脫氫酶(4-aminobutyraldehyde dehydrogenase,ABALDH)轉化為GABA[4,7-8]。
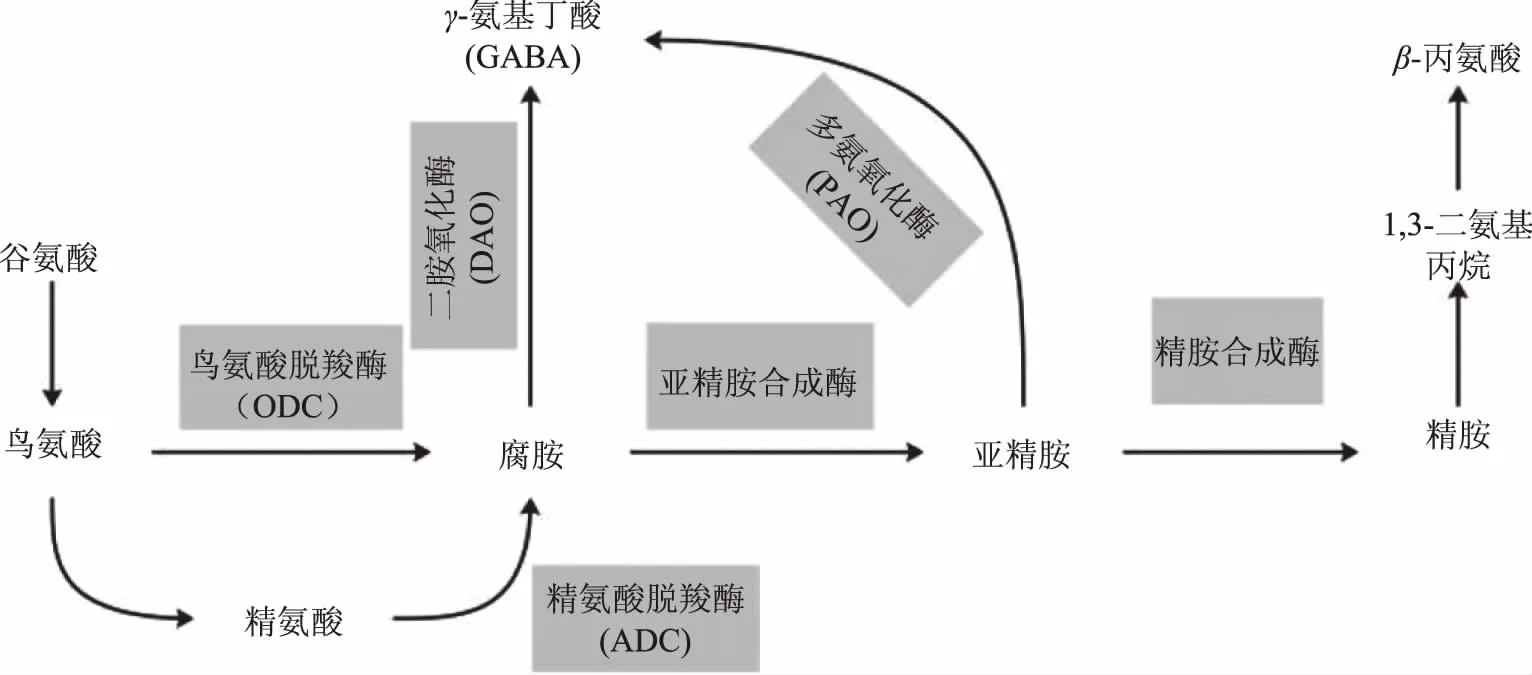
圖2 多胺降解生成GABA代謝路徑Fig.2 The production of GABA through polyamine degradation
1.3 環境脅迫促進GABA富集機理
當外界環境發生改變,不利于植物生長時,會激發植物內源酶活性,進而大量產生GABA,從而使植物適應外界的變化。GABA對于植物在逆境環境下的生存具有重要意義。
GABA在植物體內主要有以下作用:a、調節pH,當植物在無氧或受到機械損傷時,細胞質內會產生大量的H+,從而導致細胞質酸中毒,GAD在代謝的過程中需要消耗H+,減緩酸中毒,從而起到調節pH的作用[9];b、氮素儲藏,GABA代謝的過程中會消耗大量的谷氨酸,谷氨酸中的氮素被臨時轉移到GABA,通過GABA支路代謝,氮素被重新用于蛋白合成[10];c、抵御干旱:GABA會使植物氣孔減小,從而減少水分的流失,進而具有更強的抗旱能力[11];d、防止蟲害,當受到蟲害時,植物液泡中的有機酸被釋放從而激發GAD的活性,促進GABA的產生,研究發現一定量GABA能夠減緩幼蟲的成長發育、降低其存活率[12-13];e、抵抗霉變,經GABA處理后的果實,其抗霉變相關的酶的基因表達及活性會增強,提高果實的抗霉能力[14]。
1.3.1 GABA支路 GAD是GABA支路代謝的限速酶,其活性與不同物種來源、環境條件差異等有密切聯系[15-16]。GAD的活性主要由pH和鈣離子/鈣調蛋白(CAM)結合調控[17]。在受到環境刺激時(低氧、低溫、酸刺激、高鹽滲透等),植物體內H+和Ca2+會大量增加,H+和Ca2+對于GAD的活性均有促進作用。大部分GAD在酸性環境下具有較大活性,只有少部分GAD在堿性環境下能保持較大活性,微生物GAD的最適pH一般在3.8左右[18],而植物GAD的最適pH則在5.8左右[19]。
Ca2+也是影響GAD活性的重要因素,GAD是一種鈣調素協調蛋白,含有CAM結合蛋白,能夠與Ca2+結合形成Ca2+/CAM。GAD的氨基酸組成中C端含有22~25個氨基酸殘基,形成的空間結構能夠與Ca2+/CAM結合,起到激活GAD的作用[20],進而促進GABA的合成。但Ca2+僅在與CAM共同作用下才會激發GAD的活性。植物組織受到外界刺激(冷激、熱激、機械損傷、滲透壓脅迫等)時,細胞質中的Ca2+會大量增加,激活GAD,促進GABA合成。
1.3.2 多胺降解 多胺降解生成GABA的代謝酶包括ADC、ODC、DAO、PAO以及ABALDH等[21-22]。多胺降解生成GABA的過程可以分為兩個步驟:多胺的生成和多胺的降解。多胺的生成主要由ADC和ODC來調控。ADC能催化精氨酸生成鯡精氨,進而轉化為腐胺。ADC在植物界中分布廣泛,已發現的編碼基因有ADC1和ADC2[23]。ADC在所有的植物組織中均有表達,在受到冷激作用下會促進ADC1的表達,而ADC2僅在植物組織受到外界刺激(冷凍、干燥、高鹽滲透、機械損傷等)時,會促進其表達[3]。ODC是一種磷酸吡哆醛依賴酶,能夠直接催化鳥氨酸生成腐胺,主要分布于動物組織內,在植物中表達程度較低,合成的基因有ODC1和ODC2,其中ODC2僅存在于人體。ODC在逆境脅迫下活性變化較小,其主要在細胞分裂及發育的過程中發揮作用[24]。受到外界刺激時,會促進ADC和ODC的表達,提高ADC和ODC的活性,進而促進多胺的積累[25]。
多胺的降解主要受到PAO和DAO的調控。DAO和PAO可分別將腐胺和亞精胺轉化為氨基丁醛,然后經ABALDH催化生成GABA[4]。通常,DAO在植物體內的表達程度較低,但是當受到外界環境刺激時會促進DAO的表達增加其活性,將積累的多胺轉化為GABA[25]。
2 環境脅迫處理在富集GABA中的應用
2.1 低氧脅迫
低氧條件下,GABA被認為是一種信號分子,能夠調節細胞質pH,抵抗氧化性損傷[26-27]。低氧是脅迫植物富集GABA最常用的手段之一,具有操作簡單、成本低、效果明顯等優勢,在植物富集GABA中有廣泛的應用。針對不同原材料,低氧處理方式獲得GABA含量雖有較大的差異,但較未處理組,GABA含量可提高5~30倍不等。楊潤強等[28]先用pH3.5的營養液浸泡蠶豆培養36 h,隨后低氧聯合NaCl脅迫培養4 d,蠶豆GABA含量可達1.06 mg/g(Dw),較空白GABA提高了7.57倍。尹永祺等[29]以玉米為原材料,在通氣量為1.0 L/min的條件下,脅迫培養3 d,GABA含量提高了12.2倍。黃亞輝等[30]分別用液體浸泡和真空充氮的方式脅迫茶葉富集GABA,茶葉的GABA含量分別達到4.01 mg/g和2.88 mg/g,提高了14倍和30倍。除了營養液的方式進行低氧脅迫,丁俊胄等[31]用CO2和N2真空充氣的方式對糙米進行低氧脅迫,發現CO2的脅迫效果要優于N2,且脅迫介入的時期及時間長短對富集GABA的效果也有顯著差異。Ding等[32]在使用“黑糯”和“先恢”兩個高GABA水稻親本研究中,發現常規發芽72 h后GABA含量是未發芽的2.8倍和6.4倍,而在66~72 h期間經CO2低氧處理后的樣本GABA含量分別是未發芽糙米的38.5倍和37.6倍。
以往大部分的研究認為,低氧脅迫下,植物主要是通過 GABA支路的方式富集。植物在低氧脅迫下,細胞質中的H+大量增加,促進了GAD的代謝轉化。尹永祺等[33]以發芽玉米為實驗材料,檢測低氧脅迫下玉米發芽過程中GAD、DAO、AMADH的活性,發現發芽過程,GAD活性先上升后下降,始終高于對照組,而DAO總體呈下降趨勢,AMADH呈穩定趨勢。玉米發芽過程中,多胺降解酶始終處于較低的活性,而GAD則處于較高的活性,表明在該過程中是GABA支路起主要作用。但最近的研究發現由多胺代謝所產生的GABA占了相當大的比例,氨基胍對多胺氧化酶有特異性抑制作用,郭元新等[34]在低氧脅迫下用2.5 mmol的氨基胍抑制大豆中DAO的活性,而對GAD等酶無抑制作用,其富集GABA的含量降低了32%,說明大豆富集的GABA中有32%來自于多胺降解。
2.2 高鹽滲透
高鹽滲透也是富集GABA的手段之一,植物組織在培養的過程中添加高濃度鹽溶液,可以促進植物富集GABA。NaCl是高鹽脅迫中常用的脅迫劑。鹽脅迫的研究主要集中在高鹽對GABA支路的影響。鹽脅迫能夠提高植物體內GAD的活性,曾晴等[35]用不同濃度的NaCl處理大豆,發現高鹽滲透條件下,GAD活性和GABA含量都隨著NaCl濃度的增加而升高,當NaCl濃度為100 mmol/L,GABA含量可達269.93 mg/100 g。同時,鹽脅迫作用在玉米、糙米,苦蕎等植物中也能起到類似的促進作用[36-39]。鹽脅迫除了能提高GABA支路的積累量,對一些植物的多胺降解途徑也有一定程度的提高,Xing等[40]以大豆為原材料,通過鹽脅迫處理,發現隨著NaCl濃度的提高,DAO的活性增強,多胺(Put、Spd和Spm)含量降低,GABA含量提高了17倍。
2.3 低溫脅迫
低溫脅迫是指在較低的溫度下儲藏處理植物,在該過程中會增加植物組織中GABA的含量。植物為了增強其抗凍能力,通過GABA支路提供能量,減少活性氧積累[27]。低溫脅迫是一種操作簡單、高效的處理方式,一般不會單獨使用,會在植物種子發芽前后或聯合其他脅迫方式共同使用。馬麗等[41]采用低溫脅迫后的糙米進行發芽實驗,結果顯示經低溫脅迫后的發芽糙米GABA含量高,為原材料(未冷凍處理未發芽組別)的2.58倍,是發芽組(未冷凍處理)的1.19倍。白青云等[42]將馬鈴薯用營養液浸泡后,用-24 ℃和-68 ℃冷凍,隨后微波解凍,結果顯示-24 ℃處理要比-68 ℃處理GABA含量更高,GABA含量可達51.37 mg/100 g,為原材料5.61倍。低溫脅迫與其他脅迫方式聯用可以進一步增加植物中GABA的含量,尹永祺等[43]將低氧處理后的發芽玉米,分別置于5、-5、-18 ℃進行低溫脅迫處理,解凍4 h后測定GABA含量,發現在-18 ℃條件下GABA含量最高,約為原材料(未低氧未低溫脅迫處理)GABA含量的30倍,同時較、單一低氧處理GABA含量提高36%。Wang等[44]用NO處理冷藏香蕉,發現低溫儲藏條件下,NO可以增加DAO酶活性同時降低GABA-T活性,促進GABA的積累。綜上,低溫脅迫對兩種代謝途徑均有作用,可通過增加GAD活性、DAO活性或降低GABA-T活性等方式促進植物中GABA的積累。
2.4 超聲波脅迫
超聲波技術常用于提取植物天然活性成分,提高植物種子發芽率,食品的清洗滅菌等。鄭藝梅等[45]采用28、40 kHz的超聲波分別處理發芽糙米,能夠顯著提高其GABA含量,約為原材料的3倍。Yang等[46]報道了浸泡過程中超聲波處理可提高黃豆芽中的GABA含量,使用頻率為40 kHz、功率300 W超聲波處理30 min的樣品組GABA含量比對照組高出43.39%。張祎等[47]報道了在糙米發芽17 h時采用頻率30 kHz的超聲波處理16 min,GABA含量達到77.56 mg/100 g。Ding等[48]研究25 kHz超聲波處理不同發芽時段糙米中GABA積累量,發現發芽時間0、66 h的糙米,經5 min超聲處理后均可顯著提高GABA含量,分別較未超聲處理組提高60.87%和69.24%。
2.5 其他脅迫方式
除了低氧、高鹽、低溫等脅迫方式,其他方式如高溫熱激、酸性刺激等處理方式均能有效增加植物GABA的含量。Young-Su Youn等[49]采用低氧聯用熱處理方式,低氧處理后經120 ℃處理30 s,脅迫兩種小麥發芽富集GABA,結果表明聯用脅迫方式處理的,兩種小麥GABA含量較、單一低氧處理組分別提高了1.82 mg/100 g和2.23 mg/100 g。Li等[50]采用弱酸性電解水處理粟谷發芽,結果顯示弱酸性電解水可促進粟谷GABA積累,較原材料GABA含量提高21%。Hyun Soo Kim等[51]采用低氧、酸刺激等方法聯用處理糙米、米糠,均可以促進GABA的合成,相較未處理組,GABA含量提高了17倍,達523 mg/100 g。
3 結論
GABA支路和多胺降解均是植物GABA代謝的重要途徑,對植物在環境脅迫下生長起著至關重要的作用。與GABA支路途徑相比,多胺降解途徑生成的GABA含量雖然較低,但對于植物在環境脅迫條件下積累GABA研究具有重要意義。此外,不同脅迫方式對植物GABA含量及代謝機制均有較大差異,研究不同脅迫方式對植物積累GABA的影響,可用于研發高GABA食品。聯用不同脅迫方式可進一步促進GABA積累,但其涉及的不同代謝途徑及其相互作用機理尚不明確,有待進一步研究。
猜你喜歡
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
中國生殖健康(2020年6期)2020-02-01 06:28:50
新世紀智能(英語備考)(2019年12期)2020-01-13 06:07:18
中國生殖健康(2019年11期)2019-01-07 01:28:02
中國生殖健康(2018年6期)2018-11-06 07:09:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
