藥用植物四環三萜皂苷生物合成及代謝研究進展*
2018-10-30 06:23:48孟珍貴龍光強李龍根張廣輝楊生超陳軍文
世界科學技術-中醫藥現代化 2018年6期
楊 玲,范 偉,孟珍貴,龍光強,李龍根,張廣輝,楊生超,陳軍文**
(1.云南農業大學農學與生物技術學院 昆明 650201;2.云南農業大學云南省藥用植物生物學重點實驗室 昆明 650201;3.云南農業大學西南中藥材種質創新與利用國家地方聯合工程研究中心 昆明 650201)
三萜皂苷是以糖苷鍵將三萜皂苷元和糖結合從而形成的配糖體,其中包括四環三萜(達瑪烷型為主)和五環三萜(齊墩果烷型為主)兩大類,圖1所示分別為達瑪烷和齊墩果烷以及各自代表性的三萜皂苷:20(S)-原人參二醇和黃甘草皂苷。近幾十年還發現了由于氧化、環裂解、甲基轉位、重排及降解等產生的高度氧化的新骨架類型三萜皂苷。三萜皂苷結構復雜,從植物中直接提取或通過化學合成間接獲得都較為困難[1]。因此,闡明四環三萜皂苷的生物合成途徑及代謝調控對于了解三萜皂苷的整體代謝框架具有重要意義。
四環三萜皂苷(Tetracyclic triterpenoid saponins)作為三萜皂苷中重要的一類,其主要包括達瑪烷型(Dammaranes)、羊毛脂烷型(Lanostanes)、環菠蘿蜜烷/環阿屯烷型(Cycloartanes)、大戟烷型(Euphane)、葫蘆烷型(Cucurbitanes)、楝烷型(Meliacanes)、甘遂烷型(Tirucallanes)等。目前四環三萜皂苷主要存在于雙子葉植物中,靈芝屬中也有大量報道發現,如許多常用藥用植物如人參(Panax quinquefolius)[2]、黃芪(Astragalus membranaceus)[3,4]、三七(Panax pseudoginseng)[5]、麥 冬(Ophiopogon japonicus)[6,7]、升麻(Cimicifuga foetida)[8,9]、靈芝(Ganoderma lucidum)[10,11]等都含有四環三萜皂苷類成分。
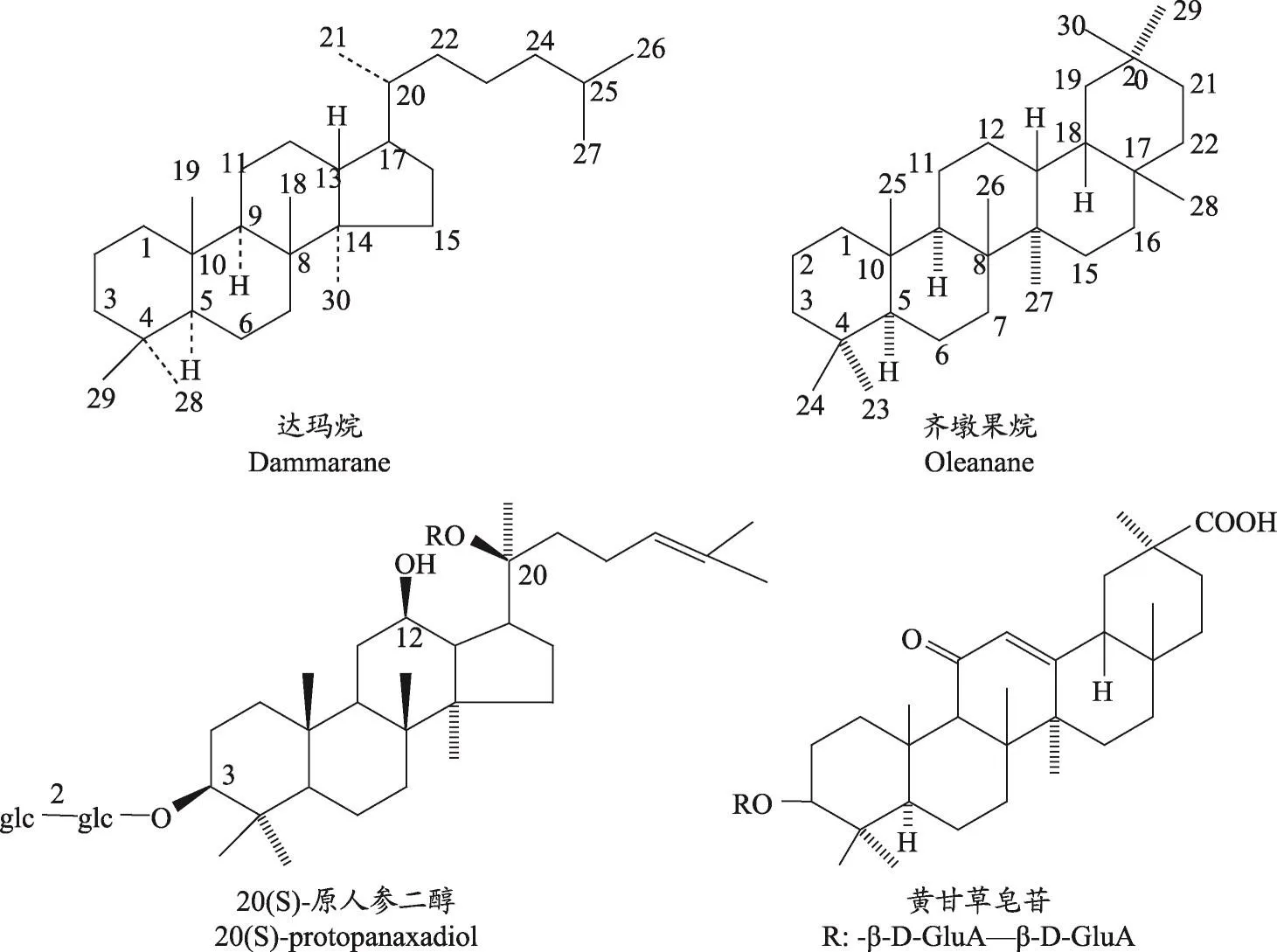
圖1 三萜皂苷中達瑪烷與齊墩果烷型皂苷結構示意圖
雖然很多藥用植物的有效成分屬于四環三萜皂苷化合物,但在藥用植物中其往往含量偏低,天然藥用植物資源十分有限,嚴重影響了其活性成分的開發利用。藥用植物資源匱乏及其種植所面臨的一系列問題推進了基因工程等技術在藥用植物領域的廣泛利用。目前三萜皂苷代謝工程研究仍處于前期初步探索過程中,對大多數四環三萜皂苷的合成途徑和代謝調控機制尚未進行深入研究,相關酶基因及其表達調控的研究報道還比較少,而通過代謝工程或基因工程進行四環三萜皂苷生物合成的工廠化生產,具有廣闊的市場前景。本文綜述了藥用植物四環三萜皂苷的一些代表性成果,探究其在次生代謝工程及基因工程領域內的研究方法與策略,以期對藥用植物四環三萜皂苷后續研究提供一定的參考價值。
1 四環三萜皂苷及其生物合成
目前研究發現,四環三萜皂苷的代謝合成主要通過甲羥戊酸途徑(Mevalonate pathway,MVA途徑)和類異戊二烯途徑(Isoprenoid Biosynthetic Pathway,MEP途徑),形成異戊烯焦磷酸(IPP)和二甲烯丙基焦磷酸(DMAPP)前體物,生成2,3-氧化鯊烯(2,3-oxidosqua?lene),再經后續的一系列關鍵酶及相關酶,如達瑪烯二醇合成酶(DS)、環阿屯合酶(CS)、羊毛甾醇合酶(LS)、細胞色素P450單加氧酶(CYP450)、糖基轉移酶(UGT)等進行三萜骨架修飾,從而合成多種結構和功能各異的四環三萜皂苷[12,13]。
1.1 達瑪烷型三萜皂苷
達瑪烷型如五加科植物人參、三七和西洋參(Panax quinquefolius)等的根、莖、葉、花、果實中均含有多種人參皂苷[14-18],酸棗(Ziziphus jujuba)仁中的酸棗仁皂苷(jujuboside)A和B[19],以及絞股藍中的絞股藍(Gy?nostemma pentaphyllum)皂苷[20,21],其苷元絕大多數屬于達瑪烷型四環三萜,達瑪烷型三萜皂苷結構從環氧鯊烯由全椅式構象形成達瑪烷骨架,主要分為20(S)-原人參二醇和20(S)-原人參三醇。達瑪烷型皂苷在心腦血管系統、抗衰老及抗腫瘤等方面具有廣泛應用[22-25],迄今為止其研究仍關注在少數幾種植物中,深入挖掘達瑪烷型皂苷生物合成及代謝機理將有助于后續對達瑪烷型皂苷的開發利用。
1.2 環阿屯烷型三萜皂苷
環阿屯烷型基本骨架在C19位甲基與C9位脫氫形成三元環,其母核結構在C13,C14位分別連接有β、α-CH3,環阿屯烷型三萜皂苷在豆科、五味子科、毛茛科、大戟科、楝科、仙茅科等植物中均有報道其化學結構及藥理活性的相關研究[26-29],如黃芪具有補氣強壯之功效,從其中分離鑒定出20個左右的皂苷,多數皂苷元為環黃芪醇[30];環阿屯烷型三萜除在中藥黃芪中發現外,在升麻族植物中廣泛存在[31];白芨(Bletilla striata)鑒定出環阿屯型三萜并且研究了其抗血管生成活性[32]。
1.3 其他四環三萜皂苷
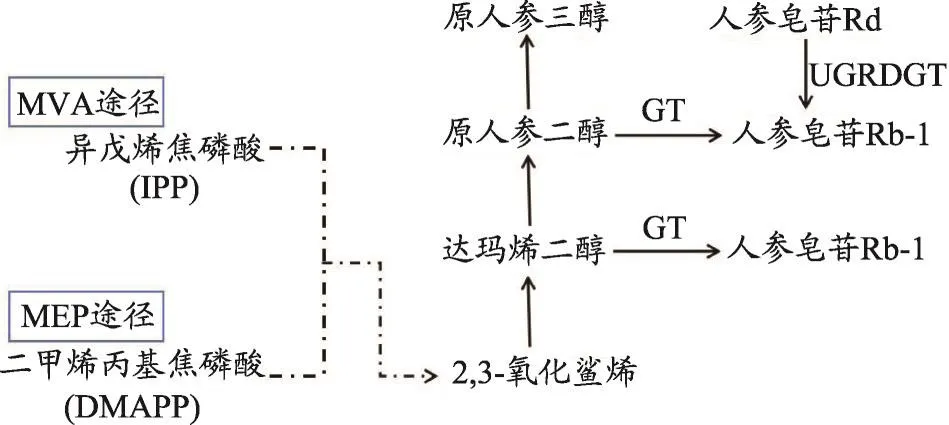
圖2 人參皂苷Rb1和Rg1生物合成途徑
羊毛脂烷亦稱羊毛脂甾烷,與達瑪烷型極為相似,不同點在于其C18甲基連在C13位,有研究表明在靈芝中分離出羊毛脂烷型三萜皂苷,是羊毛甾烷的高度氧化化合物[33-35]。此外,從葫蘆科雪膽屬雪膽(Hemsleya chinensis)中雪膽甲素、雪膽乙素均屬于葫蘆烷型[36-39];有研究發現無患子(Sapindus mukorossi)中含有大戟烷型三萜皂苷[40];楝烷型三萜皂苷主要在楝科植物中有所報道[41-43];而甘遂烷型三萜皂苷在植物分布中比較罕見,在臭椿屬(Ailanthus Desf.)中有分離得到[44-47],從生源合成來看,甘遂烷型三萜等原檸檬苦素是形成四降三萜的前體物質[48]。目前羊毛脂烷型、葫蘆烷型、大戟烷型、楝烷型、甘遂烷型三萜皂苷相關文獻報道較少,大多數研究還停留在其化學成分的提取分離鑒定及生物活性分析方面,對于生物合成與代謝調控的研究尚淺。
1.4 四環三萜皂苷生物合成
在四環三萜皂苷生物合成過程中,值得注意的是2,3-氧化鯊烯經環氧角鯊烯環化酶(OSCs)的催化發生環化反應這一關鍵步驟[49-51]。不同四環三萜化合物其立體構型是復雜多樣的,而作為多基因家族的OSCs,可使環氧角鯊烯發生環化,從而生成各類骨架的三萜化合物,然后通過細胞色素P450單加氧酶、糖基轉移酶和糖苷酶一系列相關酶的作用等對骨架進行氧化置換和糖基化等化學修飾,最終生成不同類型的四環三萜皂苷類化合物。由于受到多種因素的調節,且在這一合成的動態變化過程中所涉及到的各種中間產物和酶具有復雜性,因此目前對四環三萜皂苷的合成機理認識尚不清晰。
以典型的四環三萜皂苷——人參皂苷Rb1和Rg1為例[52]由人參皂苷Rb1和Rg1生物合成途徑(圖2),可見以,2,3-氧化鯊烯為分支點,不同的OSCs催化2,3-氧化鯊烯,獲得達瑪烯二醇(Dammaranediol),再經過糖基轉移酶的化學修飾,生成人參皂苷Rb1和Rg1[53-55]。全化學合成在人參皂苷生產中尚未得到實現,因此對那些含量少而活性強的人參皂苷單體的研究具有一定的難度。雖然目前對四環三萜皂苷生物合成途徑的研究逐漸深入,但由于其中、下游合成途徑中的多基因家族酶的結構及類型復雜多樣,導致離其合成具體步驟的闡明還有一段距離。因此,若要將生物代謝工程及基因工程的方法應用到四環三萜皂苷的合成中,還需對四環三萜皂苷類化合物生物合成過程中在碳環骨架建立后所涉及的各種復雜的官能團反應進行深入研究。
2 皂苷的代謝調控
目前有大量文獻報道關于五環三萜皂苷的合成及代謝調控,相對四環三萜皂苷研究而言,要更為深入[56]。根據四環三萜皂苷相關研究發現,在其代謝合成途徑中,一方面與外界環境因素息息相關,這些因素包括激素、誘導子和環境刺激等[57-60],而這些作用具體是通過哪種途徑實現的目前尚未研究清楚,需要進一步探索。另一方面關鍵酶基因從基因轉錄水平及轉錄后或翻譯水平上對其進行調控[61],即通過基因工程調節合成路徑中一些關鍵化合物的代謝流,利用調節基因和轉錄因子來調控關鍵酶編碼基因的表達水平,或使關鍵酶編碼基因過表達,都有可能有利于四環三萜皂苷類化合物的大量合成。在代謝途徑中酶的調節作用是極為重要的,但由于目前路徑研究尚未清楚,特別是受一些下游合成路徑中涉及的復雜酶類的影響,導致四環三萜皂苷類化合物全合成研究成為一大難題。
2.1 四環三萜皂苷的代謝調控
2.1.1 關鍵酶基因的調控
根據已推斷出的四環三萜皂苷類化合物生物合成路徑[62],人們對其中部分關鍵酶及其基因進行了深入研究。參與四環三萜皂苷生物合成路徑的關鍵酶,總體上可分為上游催化IPP和DMAPP合成的關鍵酶;中游催化IPP、DMAPP生成和萜類碳環骨架化合物以及中間體形成的關鍵酶;下游催化中間體和萜類化合物進行復雜的結構修飾從而獲得不同四環三萜皂苷的關鍵酶。近年來,眾多學者關注與研究的熱點仍與四環三萜皂苷整個生物合成途徑的闡明相關,特別是關于三萜碳環骨架合成及環上復雜官能團化等相關酶基因的鑒定與克隆,了解這些相關酶及其基因在合成途徑中的作用對四環三萜皂苷類化合物的代謝調控及更好發揮其藥用價值具有深遠意義。
迄今為止,關于四環三萜皂苷類化合物的代謝過程中上游關鍵酶研究已經比較清楚,而中下游關鍵酶對代謝的影響尚未明確。有研究表明,過量表達SS基因,可促進刺五加(Eleutherococcus senticosus)中三萜皂苷類化合物的大量合成,表明SS基因在絞股藍皂苷生物合成途徑中具有關鍵性地位[63]。蔣軍富等利用RTPCR和RACE技術,對絞股藍SE基因進行了克隆及序列分析[64]。Han等發現PgSQE1的表達受阻將導致人參皂苷的產生減少,其參與調節人參皂苷的生物合成[65]。Liu等研究2年生人參中PgSS和PgSQE1基因表達水平最高,可間接調控皂苷合成,PgDS轉錄水平與人參皂苷Rg1,Re和Rf顯著相關,PgSS基因表達與人參皂苷Rb1合成呈負相關[66]。Dhar等對催眠睡茄(Withania somnifera)進行分析確定了角鯊烯合酶、角鯊烯環氧化酶、環蒿醇合酶、細胞色素P450還原酶等在其不同發育階段的代謝產物中的表達模式[67]。缺乏相關基因的克隆及分析,是了解其生物合成、代謝調控及限制生物技術手段干預合成的主要瓶頸之一。
氧化鯊烯環化酶(OSC)是調控代謝流進入四環三萜皂苷途徑的重要分支位點,而DS是OSC家族成員之一,是合成達瑪烷型人參皂苷的關鍵靶酶,也是導致三萜產物多樣性的一個重要原因。Luo在研究中表明DS在四年生三七根部高表達,并獲得了參與三萜皂苷合成的候選基因,包括CYP450s和UGTs[68]。此外,在這些關鍵酶當中,值得注意的是P450s和GTs這兩類在植物代謝過程中占據重要地位的多基因家族,其中P450s可以利用氧氣和NAD(P)H催化三萜骨架羥基化,使其實現區域專一性及立體專一性,是四環三萜皂苷生物合成路徑中的關鍵酶[69-73]。Han等在報道中提到,原人參二醇合成酶CYP716A47屬于一個細胞色素P450單加氧酶,其參與催化達瑪烯二醇II羥基化在C12位產生原人參二醇[74]。目前,對CYP450的晶體結構以及相應的蛋白結合位點并未給出明確的說明,導致有關四環三萜皂苷生物合成途徑的下游修飾機制未能完全闡明。
2.1.2 其他調控因素
研究表明,某些激素,如茉莉酸甲酯(MeJA)等,對四環三萜皂苷合成代謝中關鍵酶基因的表達具有調節作用[75]。在人參不定根組織培養中,培養基中施加一定濃度的MeJA后,促進了人參SS基因的表達[76]。Choi等研究表明SE等三個基因與三萜皂苷合成高度相關,而MeJA通過誘導SE基因高表達從而提高三萜皂苷合成量[77]。Oktae等利用MeJA處理人參不定根,發現PgSS、PgSE和DS基因表達水平改變從而對人參皂苷Rb1、Rg1含量造成不同影響[78]。關于激素對合成代謝的調控目前尚在研究初始階段,大部分研究集中在少數幾種激素,然而不同激素對四環三萜皂苷的生物合成影響需要更進一步研究,并且激素之間的互作也要引起大家的重視。
其他外界刺激如光照、土壤水分、金屬離子、外源物質等對藥用植物中四環三萜皂苷合成的影響也逐漸引起人們重視[79-81]。例如,陽生植物對光照的需求較高,光強較大時三萜皂苷的合成較為旺盛,如絞股藍在相對光照度為70%左右時絞股藍總苷量最高[82]。光反應產物是次生代謝的必需物質,因此光照強度影響植物體三萜皂苷生物合成機理極有可能與光反應產物相關。李佳洲等研究發現土壤水分含量為0.45W時有利于人參皂苷Rg1的積累,人參皂苷Rb1則在0.70W處理下積累最多[83],可見土壤不同水分含量下四環三萜皂苷積累有所差異。此外,有研究表明,金屬鎘(Cd)脅迫誘導內源NO的產生,而NO促進SE的表達,抑制DS的表達水平,即抑制了2,3-氧化鯊烯向達瑪烷型三萜皂苷的代謝流[84]。胡向陽等利用殼聚糖處理從而提高人參皂苷合成中皂苷的積累,發現其SE轉錄水平提高[85]。由于研究尚未透徹,需要人們更進一步對于這些因素做出準確合理的分析。
四環三萜皂苷類化合物種類較多且合成途徑長而復雜,這給控制其代謝流的遺傳操作帶來極大挑戰。明確相關酶基因表達在組織和發育過程中的調控機制,對基因時空調控作用的各種順式和反式作用因子進行研究,從而進一步補充和完善四環三萜皂苷類化合物生物合成途徑及其網絡,將為今后實現大規模生產四環三萜皂苷類化合物奠定基礎。
3 結論與展望
近年來,藥用植物皂苷在醫藥領域的應用越來越廣泛,市場需求日益增高,這一現狀導致如何提高藥用植物中皂苷含量成為目前研究熱點。在藥用植物次生代謝產物生產中,制約四環三萜皂苷產量提高的原因是多方面的,但歸根結底還是由于其生物合成途徑和代謝機制尚未明確、基因工程手段還有待完善。通過對藥用植物四環三萜皂苷合成途徑的研究,進一步明確其代謝機制,從而推動藥用植物皂苷代謝工程的發展,促進藥材種植規范化、品種優良化、資源可持續化的發展。因此,未來可以從以下幾個方面深入研究。
3.1 藥用植物四環三萜皂苷類次生代謝物多樣性
藥用植物分布廣泛,而四環三萜皂苷種類多且生物合成途徑復雜,導致目前研究較為清楚的仍只是其中的很小一部分,僅鑒定和克隆了少數基因,對其代謝途徑的整體調控以及次生代謝途徑之間的協調作用等方面依舊存在很多盲區,而利用系統生物學方法開展預見性代謝工程將會拓寬未來的研究領域。
3.2 合成誘導機制多樣性
一種四環三萜皂苷類化合物的合成可能受幾種因素的誘導,一種因素又可誘導不同類型皂苷的合成積累,同一因素對不同藥用植物的誘導作用也具有差異性,這給藥用植物四環三萜皂苷在代謝調控機制方面的研究帶來極大挑戰,使得實際應用藥用植物次生代謝調控生產皂苷類化合物增加了一定難度。探究藥用植物次生代謝的具體路徑及其調控網絡,從不同側面整合與分析代謝過程及其調控手段,提高各種調控之間的聯合使用,在未來藥用植物研究中具有一定指導意義。
3.3 基因工程手段局限性
某些藥用植物繁殖周期長,在遺傳育種上難度大,不同類型的四環三萜皂苷合成可能具有單獨的合成途徑和獨特的酶促反應機制,導致基因工程手段的實施具有一定的盲目性和局限性,極大影響了研究工作的進程。未來研究中要運用各種代謝調控手段干擾代謝流,將四環三萜皂苷的化學成分研究與基因表達分析充分結合,不斷將新興技術融合應用到實驗研究中。
雖然對很多藥用植物四環三萜皂苷生物合成途徑已有了一定的研究,但由于其合成中、下游途徑中的多基因家族酶的結構及類型的復雜性,大多數合成具體步驟尚未得到清晰的闡明。隨著對藥用植物四環三萜皂苷生物合成關鍵酶基因調控研究的深入,闡明其生物合成途徑中的關鍵酶基因及其協同性,利用人工手段調控關鍵酶基因表達水平,嘗試經細胞培養、生物轉化、發根培養等一系列工業化生產去實現大規模生產藥用植物皂苷,這將是未來可能實現的一大突破。可以預見,隨著對藥用植物四環三萜皂苷合成代謝網絡的深入全面解析和相關關鍵酶基因的克隆,以及現代生物技術手段的不斷完善推進,藥用植物四環三萜皂苷代謝工程將更加有效、更好地滿足人類的需求。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
航空世界(2020年10期)2020-01-19 14:36:20
科技傳播(2019年22期)2020-01-14 03:06:54
中學生數理化·七年級數學人教版(2019年10期)2019-11-25 07:33:58
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中學生數理化·高一版(2018年9期)2018-10-09 06:46:50