五臺山林線過渡帶華北落葉松格局過程和尺度分析
2018-11-15 03:05:50賈美玉張金屯丁沫沖賈冰玉
生態學報 2018年20期
賈美玉,張金屯,*,梁 鈺,丁沫沖,曹 格,賈冰玉
1 生物多樣性與生態工程教育部重點實驗室,北京師范大學生命科學學院, 北京 100875 2 山東農業大學林學院,泰安 271000
高山林線是高山帶一個非常獨特的研究實體。高山林線的喬木處于高寒、嚴酷氣候脅迫的臨界狀態,對于全球性和區域性氣候變化的反應敏感,它對全球氣候變化具有預警作用[1- 2]。高山林線是高山帶一條重要的生態界線,是郁閉森林與高山植被之間的生態過渡帶[2]。林線過渡帶植被對氣候的響應按時間長短分為短期、中期、長期。一年左右短期的響應反應在個體特征上,幾年到幾十年的中期響應反應在生理響應上如年輪、高度和存活率等,百年以上的長期響應反應在多次演替的群落特征上[3]。林線研究的空間尺度也很豐富,多為局域尺度林線的研究[3- 4],也有跨區域尺度的研究[5-6]。對林線過渡帶的研究有研究林線樹木種群生長變化的[7- 8],有研究林線位置與氣候響應變化的[9],但在外貌、格局和動態的研究相對較少,增加以上研究內容可以探究林線群落除環境因素作用以外的主導生態過程,有研究證明緯度和海拔高度對林線樹木的限制大于人為干擾對林線的限制[3],2300m以上有樹木種群建立但它的種子密度和生長率也會很低[10]。但也有實驗結果表明草食動物對林線生態系統的限制作用也不容忽視[11]。對生長在2300m海拔以上并受放牧干擾的人工林生存變得異常艱難,它的種群年齡特征和格局過程的尺度研究也顯得尤為重要,研究結果不僅可以捕捉種群發展的特征還可以推斷類似地區生物和非生物因素對喬木的作用尺度為后期管理提供參考。
五臺山屬于溫帶落葉闊葉林區,是典型高山林線山地,林線附近的喬木為人工種植的華北落葉松[12]。五臺山亞高山草甸是華北地區最典型,類型最豐富,草質和生產力最高的山地草甸,對亞高山草甸的研究包括合理利用、生態評價[13-14]以及群落生態學數量分析[15]。如五臺山高山草甸群落分析表明地形因素影響群落型及優勢種的劃分,同時多樣性也是影響劃分的關鍵因素[16],五臺山草甸中小格局分析揭示格局尺度對草本群落的生態意義[17]。作為著名風景名勝區,干擾的研究集中在旅游和放牧兩方面,DCA方法分析了旅游干擾對五臺山草甸的影響[18]。放牧對高山、亞高山草甸植被-土壤系統的影響[19],對牧草品質的影響表明短期的中度放牧對牧草品質有利[20-21]。五臺山人工華北落葉松林是重要的景觀組成,林線的華北落葉松存活狀況堪憂,而它的存活率對于草甸的生產力和多樣性影響很大,它的格局對于林下草本入侵也至關重要。已有研究利用聚類和排序分析了五臺山高山林線的植被景觀,得出了陰坡和陽坡的林線的海拔范圍和草本植被群落類型分布的海拔范圍[22],野外儀器測定分析五臺山林線喬木的光合作用對生境差異的響應得出光照和溫度相比水分是光合作用更為重要的限制因子[23]。本文在前面研究的基礎上,希望通過點格局方法進一步分析華北落葉松的種群特點以及存活影響因素。點格局分析方法可以研究多尺度內森林種群的分布、生態過程對其格局的影響以及種群存活和死亡的動態[24],因此是有效的格局分析方法。
本實驗選擇五臺山南臺母樹林基地樹齡30以上的林線過渡帶為研究樣地,對樣地內的華北落葉松種群進行分析。通過分析希望探究①林線過渡帶華北落葉松種群生長特征以及死亡率與胸徑分布的關系;②不同尺度內華北落葉松活立木和枯立木分布格局,以及空間異質性和種群密度對華北落葉松枯立木和活立木格局分析,進而推測其死亡的主要原因。
1 研究區概況
五臺山位于山西省東北部,38°27′—39°15′N,112°48′—113°50′E。五臺山位于蒙古高壓中心的東南邊緣地帶。西伯利亞干冷氣流吹向海洋的過程中,以強度波動性經過五臺山向東南推移。海拔2250m以上山地的年均溫約為-1.6—-4.9℃,年降水量717—840mm[25]。五臺山所處地區屬暖溫帶半濕潤季風氣候。五臺山地區地形高聳,又臨近華北平原,是接受東南海洋夏季風送來的暖濕氣團較早和最佳的場所,五臺山山頂年降水量高達900mm以上。五臺豆村日照時數年平均2684.2h,日照充足[26]。由于地形變化較大,氣候條件差異明顯。植物資源豐富,擁有華北地區面積最大的亞高山草甸[27]。調查樣地在2300m左右,屬于山地草甸土。
2 材料與方法
2.1 野外取樣
2017年7月初在五臺山南臺母樹林基地以上選擇有顯著林線森林分布的高坡(2315m),坡向北偏西25°,坡度26°。從有顯著林草交錯帶界限為起始的一邊,往下坡方向延伸設置50m×50m的樣地,用1m為單位刻度的100m長的樣繩把樣地分成5條樣帶,每條樣帶5個樣方,共25個10m×10m的樣方,對25個樣方進行編號,在每個10m×10m樣方中對喬木個體的位置和胸徑信息進行調查,選定原點,在樣地內記錄每一棵華北落葉松(死+活)的位置和胸徑信息。在50m×50m的樣地內隨機設置5個10m×10m的樣方進行群落物種組成信息調查,統計出現的喬、灌、草物種。
2.2 數據處理與分析
點格局分析和零模型模擬的方法可以用來探索種群格局、種內兩種不同屬性個體間關系、種間關系以及生態過程在不同尺度上的效果。本研究選取成對相關函數g(r) 值為縱坐標,個體間距離為尺度作為橫坐標進行繪圖。首先運用完全空間隨機CSR(Complete spatial randomness)模型對樣地內華北落葉松的活立木和枯立木分別進行多尺度上的格局判斷,此處結果可以比較多個尺度上活立木和枯立木分布特征;再者異質性泊松過程HP(Heterogeneous Poisson process)模型模擬樣地內的空間異質性對格局的影響,通過異質性模型模擬結果的檢驗可以了解空間異質性對華北落葉松格局的作用是否顯著;最后運用隨機標簽RL(Random Labeling)模型對活立木、枯立木個體進行尺度上的相關性分析,我們可以得到它們的相關關系以及進一步推測華北落葉松死亡原因。Monte-Carlo 方法用于檢驗實際值是否顯著偏離零模型,一個點進行199個次模擬,在199次模擬中計算第五個最低值和最高值來獲得α=0.05 的顯著水平的值。在CSR模型進行格局判斷結果中,當實際值超出置信區間的上包跡線為聚集分布,超出下包跡線為均勻分布。GoF test(Goodness of fit test)可以用于檢驗模型描述實際格局值的能力。比較選定范圍內觀察值和理論值的偏差,用秩檢驗判斷顯著性,當觀察值的秩(rank)大于190說明觀察顯著偏離理論值。說明在α=0.05 水平上實際值顯著偏離模型。模型不能很好解釋形成格局的原因[28]。
K函數和g函數公式如下[29]:
式中,A為樣方面積,n為研究個體的總點數,r為兩點之間的距離即尺度,uij是i和j之間的距離,當uij
g(r)=dK(r)/dr×2πr
式中,g(r)>1,聚集分布;g(r)<1,均勻分布;g(r)=1,隨機分布。
以下為3個零假設模型的介紹[28,30]:
a.完全空間隨機CSR(Complete spatial randomness)模型
完全空間隨機模型是最簡單、常用的零模型,它假設個體之間不存在相互作用。它也叫同質性泊松分布,即研究區內所研究個體的密度是恒定的,且所研究個體在研究區域內的分布符合泊松分布,研究區域內每個點出現研究個體的概率是相同的。完全空間隨機模型可以描述種群的真實格局,但對格局形成的生態過程缺乏解釋。
b.異質性泊松過程HP(Heterogeneous Poisson process)模型
空間異質性常存在于研究區內,這時我們需要運用異質性泊松過程考慮樣方內空間一次特征的零模型假設,進而模擬植物空間分布的二次特征。在異質性泊松過程中,所研究個體在研究區域內的密度是不恒定的,而隨位置變化而變化,由λ(x,y)表示密度,其是運用圓形動窗在樣方內進行插值運算而估計出的值,從而代表空間的一次特征,計算公式為:
式中,C(x,y)(R)是以(x,y)為圓心,R為半徑的圓形動窗,Point[X]表示區域X內的點數,Area[X]表示區域X的面積。在這個模型中R需要謹慎考慮,R過大結果會接近CSR模型,R過小接近真實的分布,失去了空間一次特征假設的意義。經過10、5、3m的嘗試后決定R=3m最為合適。此模型用于分析華北落葉松死亡的原因是不是與生境相關。因此采用華北落葉活立木做為背景,零模型假定活立木與枯立木的生境沒有顯著性差異。
c.隨機標簽(Random labeling)模型
隨機標簽模型屬于雙變量假設模型的一種,它可以用來分析兩個物種或者同一物種兩類型的格局在研究尺度上的相關關系。它假設兩個變量的空間分布格局由相同的隨機過程(或受到相同的限制條件)產生的。每一次循環模擬中,給變量1的點隨機打上標簽,計算其周圍研究尺度內出現變量1或變量2的個體的概率。此模型用于分析活立木與枯立木的相關關系以及枯立木形成原因。
運用Programita 軟件進行點格局分析,分析結果在Origin 9.1軟件中對g(r) 、模擬值和置信區間在0—25m上進行作圖。
3 結果與分析
3.1 華北落葉松群落結構與物種組成
調查50m×50m樣地內,共433棵華北落葉松,其中活立木366棵,枯立木67棵。根據計算可知平均密度為5.77m2/棵,所有活樹的平均高度為4.96m,枝下高為2.84m。其中郁閉度接近80%,群落中木本植物物種豐富度不高但草本植物種類多樣,灌木有華北覆盆子(Rubusidaeus)、瘤糖茶藨子(Ribeshimalense),草本植物有多葉早熟禾(Poaplurifolia)、胭脂花(Primulamaximowiczii)、紫苞風毛菊(Saussureapurpurascens)、瞿麥(Dianthussuperbus)、異燕麥(Helictotrichonschellianum)、黑柴胡(Bupleurumsmithii)、小紅菊(Dendranthemachanetii)、疏齒銀蓮花(Anemoneobtusiloba)、高原毛茛(Ranunculustanguticus)、早春苔草(Carexsubpediformis)、高山紫菀(Asteralpinus)、三葉委陵菜(Potentillafreyniana)、雪白委陵菜(Potentillanivea)、互葉金腰子(Chrysospleniumalternifolium)、峨參(Anthriscussylvestris)、紫花碎米薺(Cardaminetangutorum)、珠芽蓼(Polygonumviviparum)、北京假報春(Cortusamatthioli)等。
3.2 華北落葉松胸徑分析
華北落葉松總體的均值(11.03cm)和活立木個體的均值(11.54cm)相差很小,枯立木個體的均值(8.27cm)與活立木以及總體相差很大(圖1)。把華北落葉松不同胸徑根據大小依次劃分為5個等級,計算不同胸徑區間內個體數占總體數的比例,結果表明:活立木個體在胸徑小于10cm內數量最多,而且從圖2線圖和柱狀圖都可以看出從胸徑值小的區間到胸徑值大的區間活立木呈現顯著的遞減趨勢。枯立木個體胸徑只在小于15cm范圍內分布,但在胸徑小于10cm內分布較多(圖2)。
3.3 華北落葉松的完全空間隨機模型和空間異質性過程模型模擬分析
利用完全空間隨機模型對0—25m尺度上的華北落葉松格局進行分析發現:活立木個體在0.5、2.5m和21m上大于上包跡線為聚集分布,在14.5cm上小于下包跡線為均勻分布;枯立木個體在0—15.5m上大于上包跡線為聚集分布,22.5m上小于下包跡線為均勻分布(圖3)。
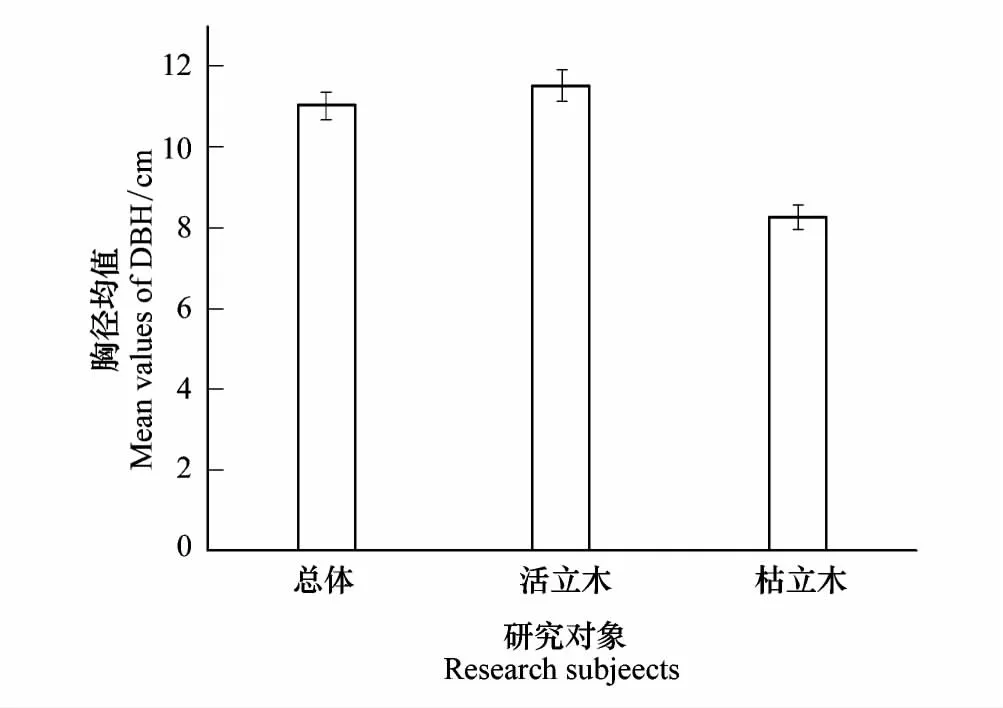
圖1 華北落葉松胸徑均值Fig.1 Average DBH of Larix principis-rupprechtii
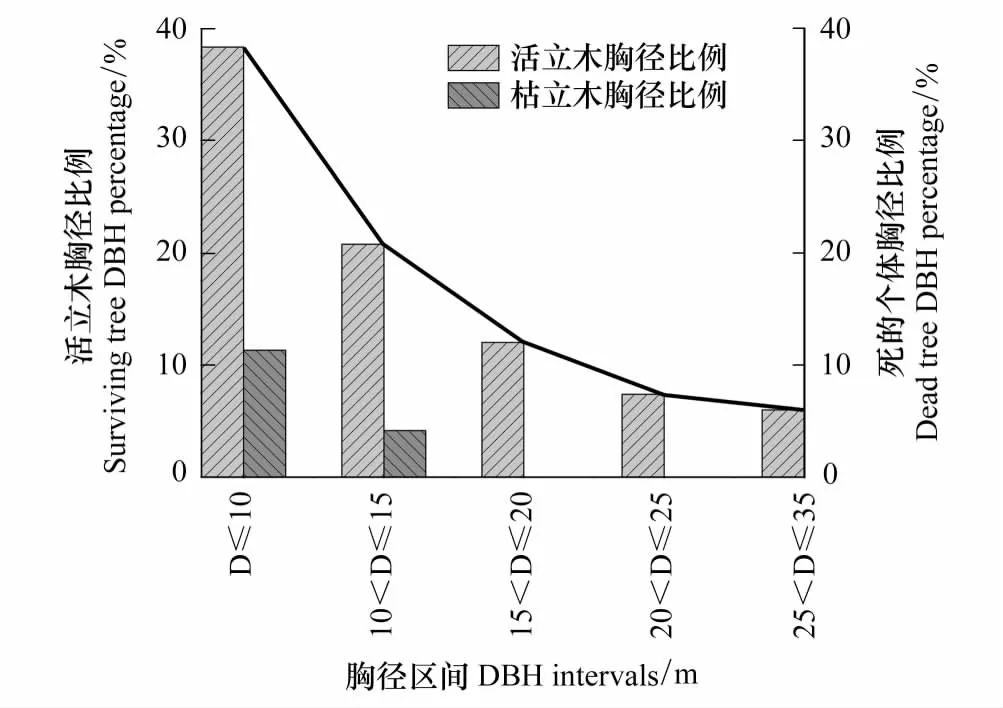
圖2 活立木和枯立木的胸徑比例Fig.2 DBH ratio of survival and dead tree
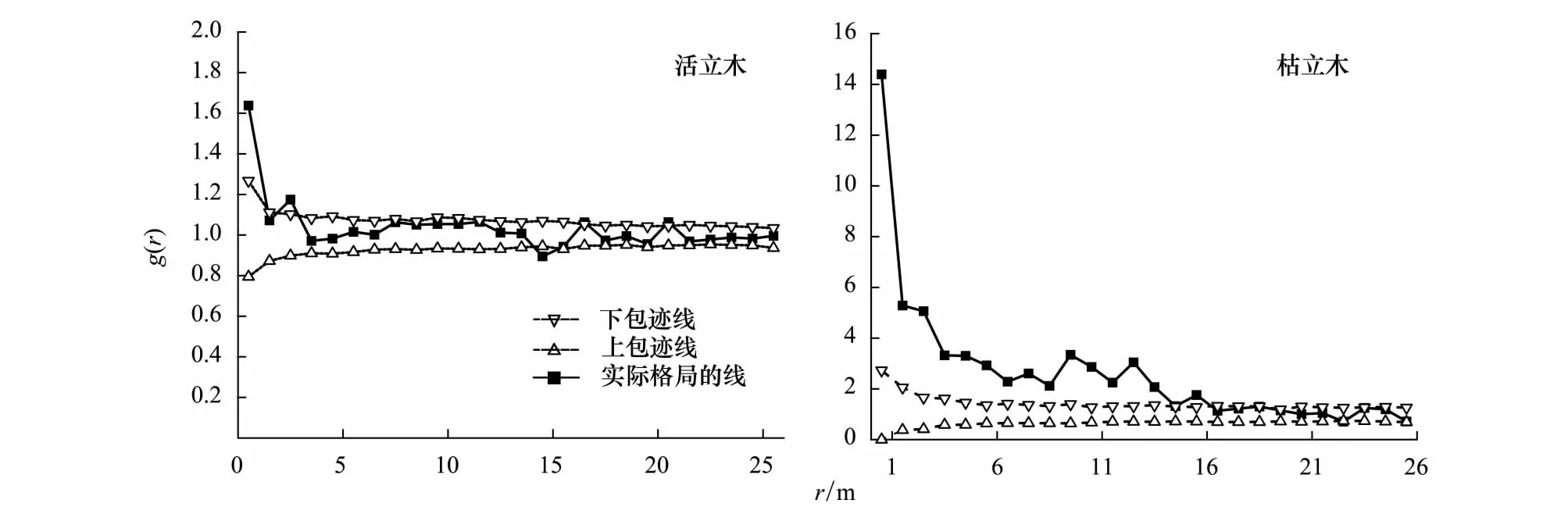
圖3 華北落葉松活立木和枯立木的格局與尺度分析Fig.3 Pattern on scales analysis of survival and dead Larix principis-rupprechtii
以華北落葉松活立木的分布密度作為空間異質性模型的一次特征值,選擇帶寬3m,Kernel方法,在0—25m上的空間異質性模擬結果表明:活立木個體格局值大部分落在空間異質性過程模型的包跡線內且HP模擬值曲線與實際格局值曲線走勢相似,空間異質性模型可以通過顯著水平95%檢驗(P=0.065)很好模擬活華北落葉松個體格局。枯立木個體的格局大部分值落在空間異質性過程模型的包跡線外且HP模擬值的曲線與實際格局值曲線相差很大,實際格局與空間異質性過程模型模擬格局的差異檢驗結果的概率值(P=0.005)小于0.05,說明枯立木格局不能被現在活立木分布為參考搭建的空間異質性模型的一次特征值的空間異質性過程模型所解釋(圖4)。
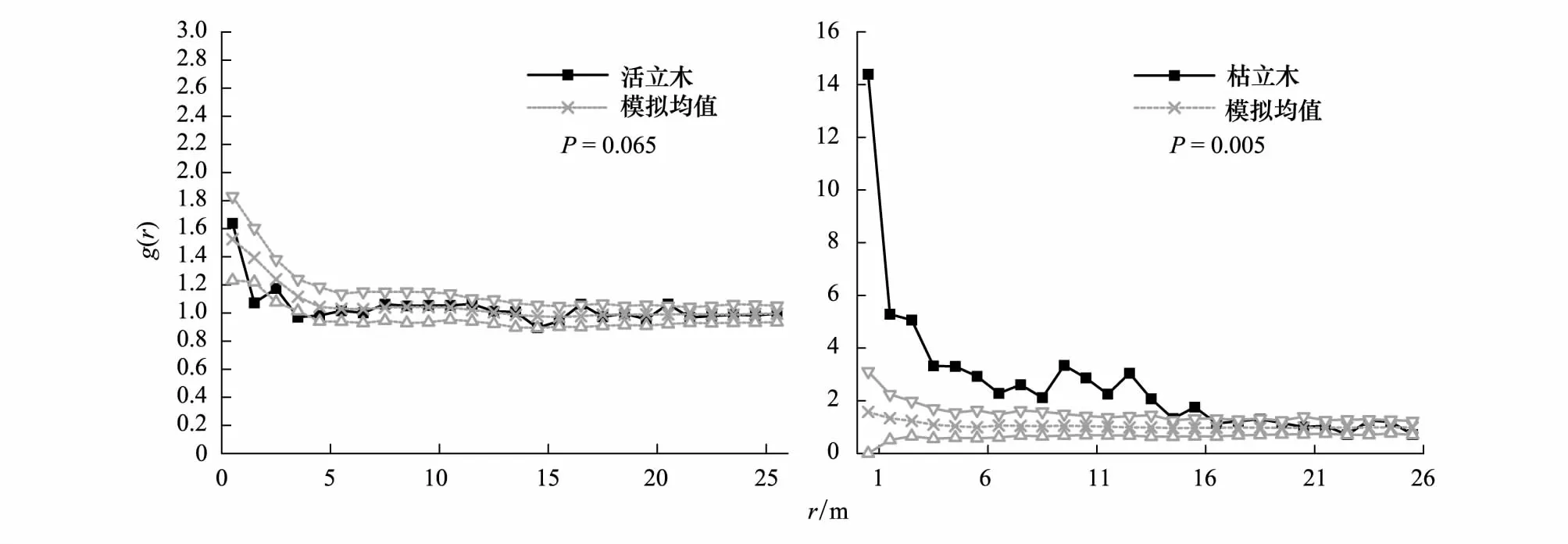
圖4 空間異質性過程模型模擬實際格局Fig.4 Heterogeneous Poisson process simulated observed patternHP 指空間異質性過程(Heterogeneous Poisson process)
3.4 隨機標簽(Random labeling)模型對華北落葉松枯立木、活立木格局的分析
采用隨機標簽對活的華北落葉松個體和死的華北落葉松個體進行分析,成對相關函數g12除在21m附近出現負相關外,在0—25m尺度上都在包跡線內,說明活立木個體與枯立木個體大部分距離內無顯著相關關系,在一定距離(21m)處才會出現負相關。g21-g11在0—15m尺度上小于下包跡線,在活立木個體附近的枯立木個體的鄰體密度低于枯立木個體附近的枯立木個體的鄰體密度。g12-g22在3m處大于上包跡線,表示枯立木個體周圍的活立木個體的鄰體密度高于在活立木個體周圍的活立木個體的鄰體密度;21m處小于下包跡線,表示在枯立木個體周圍的活立木個體的鄰體密度低于在活立木個體周圍的活立木個體的鄰體密度,其他尺度上差異不顯著。g1,1+2-g2, 1+2在0—12m尺度上大于上包跡線,枯立木個體周圍的華北落葉松(死+活)鄰體密度高于活立木個體周圍華北落葉松(死+活)鄰體密度,華北落葉松枯立木周圍密度高于活立木周圍的密度,0—15m上枯立木聚集分布,因此0—12m內高密度的華北落葉松在枯立木周圍是華北落葉松死亡的原因(圖5)。
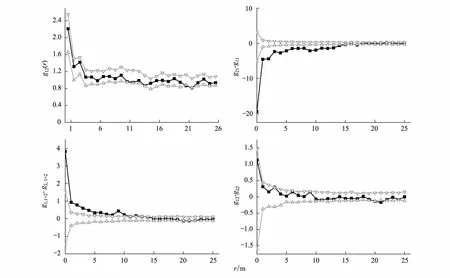
圖5 隨機標簽模型分析0—25m尺度上活立木、枯立木的相關關系Fig.5 Random labeling simulated relation of survival and dead tree on 0—25m scales“1 ” 代表枯立木 dead individuals,“2 ”代表活立木 surviving individuals
4 討論
4.1 林線過渡帶內華北落葉松種群特征
華北落葉松在研究樣地內的平均胸徑為11.54cm,而參考相關對華北落葉松人工林生長規律的研究表明,華北落葉松胸徑年平均年生長量為4.6cm[31],研究樣地內樹齡大于30年,計算得到現在它的胸徑至少應該是13.8cm,可見林線過渡帶內的華北落葉松生長速度變慢。而且隨胸徑徑級的增加活立木的數量越來越少,這說明隨著年齡的增加林線內的華北落葉松胸徑生長變慢也是其林線環境做出的適應特點之一。枯立木的胸徑在8.27cm,說明不是最近幾年的死亡,枯立木的胸徑分布在小于15cm內尤其在小于10cm內比例最高,而華北落葉松成熟年齡在43年左右,說明幼苗變幼樹的過程中死亡率最高,這與青藏高原東緣林線喬木種群生長特征有相似的結果[32]。
4.2 林線過渡帶內華北落葉松活立木、枯立木的格局與尺度的響應
針對0—25m個體距離為研究尺度,我們發現活立木在3m內有顯著聚集分布,華北落葉松冠幅比較大,猜測為幼樹在這個尺度內聚集分布,這樣可以受到成樹的庇護[33],這是適應惡劣環境的對策之一[34],在21m上也有聚集分布,可能是成樹的聚集尺度。在15m附近有均勻分布出現,可能成樹之間在這個尺度上更容易生存。枯立木的聚集分布很明顯,在15m以內都是聚集的,顯然死亡不是隨機的,這與青藏高原林線內冷杉格局研究結果支持的隨機死亡假說不一致[35]。利用活立木分布模擬的空間異質性研究結果表明,環境異質性對活立木格局形成有顯著影響[36-37],環境異質性模擬的格局與枯立木的格局不吻合,說明死亡主要不是現有的空間異質性導致的,在林線這種敏感的環境中任何微小的環境改變都會導致樹木的死亡[38],過去的空間異質性可能是導致華北落葉松死亡的原因。尤其是氣候變動大的年份等,這需要結合氣候數據進行進一步分析驗證。
4.3 隨機標簽對華北落葉松死亡的生態學推測
對活立木和枯立木的相關性在0—25m尺度上分析表明死的華北落葉松與活的華北落葉松在21m尺度附近有顯著的負相關,說明現有活立木對枯立木的種內排斥作用出現在21m附近。在0—15m上活立木個體周圍的枯立木比枯立木周圍的枯立木出現概率低,說明該尺度范圍內枯立木周圍的死亡率大于活立木周圍的死亡率,即在這個尺度范圍內華北落葉松的死亡個體很集中,但在15m內現有活立木不對枯立木產生排斥作用,推測現有枯立木出現的原因和現有活立木的分布無關。21m處活立木本身是聚集格局而枯立木自身是隨機格局,因此大于枯立木周圍的活立木,3m處活立木和枯立木本身格局都是聚集格局,但是枯立木周圍的活立木出現概率卻大于活立木周圍的活立木的出現概率,而0—15m上(含3m)活立木周圍的枯立木出現概率低,綜合二者結論說明枯立木被活立木所包圍,活立木密度在3m距離尺度內大于枯立木的密度。說明活的華北落葉松在林線周圍聚集分布且聚集強度在3m內大于死的華北落葉松的聚集分布,推測抱團生活更適應林線生境。最后對枯立木周圍華北落葉松的總體與活立木周圍華北落葉松總體出現概率的比較表明總體向枯立木方向聚攏發展,而且枯立木聚集分布范圍也很廣,推測南臺林線的華北落葉松存在負密度制約效應這與最近對森林研究的結論一致[39-40],即華北落葉松密度越高存活率降低,后期死亡風險越大。相關研究表明:幼樹階段的負密度制約效果明顯,環境異質性對灌木的幼苗作用有限[41]。根據負密度制約假說分析[41,42],上面分析推測種內排斥在0—15m內不是枯立木出現的原因,而作為灌木的華北覆盆子(Rubusidaeus)、瘤糖茶藨子(Ribeshimalense)在15m內對華北落葉松的幼苗或者幼樹的種間競爭可能是死亡率增大的原因,也有相關研究提到林線內鬼箭錦雞兒(Caraganajubata)的蓋度通常在40%以上,有時甚至達到90%,可能影響了喬木的生長[22]。因此灌木對喬木格局的影響有待研究驗證。
5 結論
林線過渡帶生境內的華北落葉松生長緩慢,死的個體胸徑都集中在幼苗到幼樹階段,活立木胸徑隨徑級增加也呈現遞減趨勢。死亡個體聚集分布在0—15m上顯著,活的個體僅在3m以內和21m處聚集分布,空間異質性對活的華北落葉松有顯著作用,但是幼樹階段的華北落葉松的負密度制約作用顯著,在現有調查尺度內由華北落葉松的活立木構建的空間異質性不能很好解釋華北落葉松的枯立木的格局,但不同林線類型的存在本身就說明大尺度上空間異質性對林線樹木的死亡起很大作用。負密度制約效應對死亡率的影響很大,種內排斥作用在近距離內不明顯。本文主要針對華北落葉松為對象展開討論,針對灌木對幼苗和幼樹成活的影響有待研究證實。
