甘薯TPS基因家族分析及IbTPS7a基因克隆與功能研究
2018-11-28 09:04:18邵歡歡鄭海燕廖若星費雪婷馬沁沁袁向華王曉燕
食品與生物技術學報 2018年10期
邵歡歡, 鄭海燕, 廖若星, 徐 攀, 費雪婷,雍 彬*, 馬沁沁, 袁向華, 王曉燕
(1.四川師范大學 生命科學學院,四川 成都 610101;2.四川師范大學 附屬中學,四川 成都 610101)
關鍵字:甘薯;海藻糖;抗逆;數字表達譜
農作物在生長過程中容易受到干旱、極端溫度、高鹽及病蟲害等脅迫,使其生長發育受到限制,進而影響產量。為了應對來自環境中的非生物或生物脅迫,經過長期的進化,植物形成了復雜的響應機制,例如植物可以改變其形態和生理生化狀態來適應環境的變化[1]。海藻糖(Trehalose)是一種非還原性二糖,由兩分子葡萄糖通過半縮醛羥基縮合反應形成的α,α-1,1糖苷鍵連接而成,在細菌、真菌、昆蟲、無脊椎動物和植物中廣泛存在[2]。海藻糖在酸、堿、高溫等條件下具有很強的穩定性和強吸水性,可以有效地保護細胞并使其具有抗脫水作用。海藻糖分子中具有較多的羥基可以與多種生物大分子結合形成氫鍵,對生物大分子能起到保護作用并降低失活風險,有助于維持細胞或生物體的正常生命過程,進而提升生物體的抗逆能力[3]。自然界中已經發現了5個海藻糖合成通路,分別是TPS-TPP途徑、TreP途徑、TreS途徑、TreY-TreZ途徑和TreT途徑,其中TPS-TPP途徑是高等植物唯一的海藻糖合成途徑[4-6]。TPS-TPP途徑是由兩個關鍵酶經過兩步反應形成,首先在TPS催化下尿苷二磷酸葡萄糖(UDPG)和 6-磷酸-葡萄糖(G6P)反應生成 6-磷酸海藻糖(T6P),T6P在磷酸海藻糖磷酸酯酶(TPP)催化下最終形成海藻糖[7]。
甘 薯 (Ipomoea batatas (L.) Lam.), 旋 花 科(Convolvulaceae)番薯屬(Ipomoea)一年生草本植物,淀粉含量高且營養豐富,是一種重要的糧食作物。中國甘薯資源十分豐富,種植面積和產量均居世界第一[8]。甘薯多被種植在不發達地區的邊緣土地中,產量很高但并不占用良田,對環境要求較低,容易受到干旱、鹽堿、極端溫度等非生物脅迫,因此其抗脅迫能力也較強。
在高等植物中已經發現了較多的TPS基因,有些植物中具有由一系列TPS基因組成的TPS基因家族,例如擬南芥基因組中具有11個TPS基因(AtTPS1-11)組成的AtTPS基因家族[9],根據擬南芥TPS基因家族序列特點在楊樹和水稻等植物中也發現了類似的TPS基因家族,其中楊樹TPS基因家族有12個PtTPS基因,水稻TPS基因家族有11個OsTPS基因[10-11]。目前還沒有關于甘薯TPS基因家族及功能研究的報道,作者根據NCBI數據庫中擬南芥和楊樹的TPS基因家族序列,利用本地Blast(BlastN和BlastP)在甘薯轉錄組數據庫[12]中共發現了7個TPS基因,成功克隆得到了IbTPS7a基因,分析了IbTPS7a基因在甘薯不同組織中的表達量,在大腸桿菌和釀酒酵母對IbTPS7a基因的抗逆功能進行了驗證,有助于了解甘薯的TPS基因抗逆功能和作用機制,為未來甘薯的育種及基因工程改造研究打下了基礎。
1 材料與方法
1.1 材料
1.1.1 薯材、菌株、質粒 甘薯品種為徐薯18,大腸桿菌克隆及表達菌株分別為Escherichia coli DH5α和 E.coli BL21 (DE3), 酵 母 表 達 菌 株 為Saccharomyces cerevisiae INVSc1,大腸桿菌表達載體為 pET-32a(+),酵母表達載體為 pYES2。
1.1.2 主要試劑、工具酶和分析軟件 RNA提取試劑 盒 MiniBEST Plant RNA Extraction Kit: 購 自TaKaRa公司;PCR高保真酶PrimeSTAR?HS(Premix):購自TaKaRa公司;酵母基因組提取試劑盒:購自北京天根生物科技有限公司;膠回收試劑盒E.Z.N.A.TMPlasmid Mini Kit I:購自Omega Bio-tek 公司;M-MLV逆轉錄酶: 購自 Invitrogen;RNase inhibitor及各種常規限制內切酶:購自Thermo Scientific公司;引物合成及測序:成都擎科梓熙生物技術有限公司合成。
1.2 方法
1.2.1 甘薯TPS基因家族分析及進化樹構建 利用已知的單子葉植物水稻和雙子葉植物擬南芥中TPS基因家族核酸序列和蛋白序列,用BlastParser軟件對甘薯轉錄組數據庫分別進行本地BlastN和BlastP(e-value為1e-10)比對,獲得相關同源序列,結合兩種比對結果分析甘薯轉錄組數據庫中可能存在的TPS基因家族。將獲得的甘薯TPS基因家族序列與擬南芥的相關序列使用MEGA 5.0軟件采用Neighbor-joining法制作系統進化樹。
1.2.2 總RNA提取及反轉錄 根據TaKaRa公司植物總RNA提取試劑盒的推薦方法對分別對徐薯18的葉片、塊根和莖提取總RNA,將去除基因組DNA后無降解的總RNA使用Thermo公司的Nanodrop 2000進行RNA濃度測定,并根據濃度按照比例將3個組織樣品的總RNA進行等量混合,以M-MLV反轉錄酶反轉錄得到cDNA第一鏈,分裝后保存在-20℃冰箱中備用。
1.2.3 IbTPS7a基因克隆及表達載體構建 以甘薯轉錄組測序數據及前期經比對得到的甘薯TPS基因家族中的IbTPS7a序列為模板,使用SLIC(sequence and ligation independent cloning) 法設計引 物 , 引 物 序 列 所 下 所 示 :TPS-32sF:5’ -GAAAACCTGTACTTCCAGGGTATGATGTCCAAATC GTATACCAACTT-3’ ;TPS-32sR:5’-GTTTAGAGG CCCCAAGGGGTTATCAATCCGAGCTTCCGGTGG-3’ ;PET_TPS-F:5’-CCACCGGAAGCTCGGATTGA TAACCCCTTGGGGCCTCTAAAC-3’ ;PET_TPS-R:5’-ACCCTGGAAGTACAGGTTTTCACCAGAAGAAT GATGATGATGATGG-3’其中下劃線部分為pET32a(+)載體骨架部分。以反轉錄得到的cDNA和pET32a(+)質粒為模板,分別以 TPS-32sF和TPS-32sR以及PET_TPS-F和PET_TPS-R為引物,使用PrimeSTAR?HS(Premix)分別進行 PCR擴增IbTPS7a基因和線性 pET32a(+)載體,PCR產物經電泳后膠回收,回收產物經T4 DNA Polymerase處理、退火,構建得到重組質粒pET32-IbTPS。把得到的pET32-IbTPS質粒經PCR驗證后送至成都擎科梓熙生物技術有限公司進行測序,以測序正確的質粒為模板使用引物TPS-yF(5’-CGGGATCCCGATG ATGTCCAAATCGTATACCAACTT-3’) 和 TPS-yR(5’ -GCTCTAGAGCTCAATCCGAGCTTCCGGTGG-3’)進行PCR,將純化后的PCR產物和pYES2載體使用BamH I和Xba I進行雙酶切,膠回收使用T4 DNA ligase于18℃過夜連接后轉化大腸桿菌DH5α,獲得的陽性克隆進行培養后提取質粒,將質粒酶切和PCR驗證后送至公司進行測序。
1.2.4 生物信息學分析 根據測序得到IbTPS7a基因 序 列 , 用 BLAST(http://blast.ncbi.nlm.nih.gov/Blast.cgi)和DNAMAN進行基因及蛋白同源序列比對;使用 ProtParam(http://web.expasy.org/protparam/)對IbTPS7a蛋白質理化性質進行預測;進化樹使用MEGA 5.0軟件的Neighbor-Joining法構建,蛋白質氨基酸進化距離計算采用Poisson correction模型,bootstrap自檢重復1 000次;信號肽使用Signal IP 4.1 Server(http://www.cbs.dtu.dk/services/SignalP/)進行預測;跨膜結構分析使用TMpred(http://www.ch.embnet.org/software/TMPRED_form.html); 亞細胞定位 使 用 Wolf psort (http://www.genscript.com/wolfpsort.html)。
1.2.5 原核表達及非生物脅迫耐受性分析 將成功構建的pET32-IbTPS質粒轉化E.coli BL21(DE3)菌株,使用含有氨芐青霉素(Ampicillin,50 μg/mL)抗性平板篩選,挑取單菌落至2 mL的液體LB中過夜培養,取500 μL菌液加入50 mL液體LB中,37℃ 培養至 A600nm為 0.4~0.6時加入 IPTG(Isopropyl β-D-Thiogalactoside,終濃度為 1 mmol/L),將50 mL菌液放置于18℃培養箱中誘導表達16 h,SDS-PAGE電泳檢測IbTPS7a蛋白表達效果。
挑取含有pET32-IbTPS質粒的陽性克隆菌落于含50 μg/mL Amp的 2 μL LB液體培養基中 37℃震蕩過夜培養,取100 μL將其接入10 mL液體LB(含 50 μg/mL Amp)中當 A600nm為 0.6 時,加入IPTG至終濃度為1 mmol/L,18℃誘導培養4 h。按1%比例,取500 μL此培養物分別移入50 mL含0.6 mol/L NaCl、0.8 mol/L NaCl、 質 量 分 數 20%PEG6000和30%PEG6000的液體LB培養基(IPTG終濃度為1 mmol/L)中,在37℃培養箱中持續震蕩培養3 d,每間隔4 h取樣并測定其A600nm值,繪制生長曲線,實驗均重復3次,含空載體的對照菌株按照相同的方法繪制生長曲線。
1.2.6 釀酒酵母轉化及非生物脅迫耐受性分析利用醋酸鋰法將pYES2-IbTPS和pYES2質粒分別轉化S.cerevisiae INVSc1,將兩種菌落接種至液體培養基中,使用酵母基因組提取試劑盒提取含有質粒的釀酒酵母總DNA,用引物TPS-yF和TPS-yR以提取得到的DNA為模板進行PCR擴增,檢測成功轉化質粒的菌株,鑒定正確的菌液劃線固體平板保存至4℃冰箱中短暫保存。
分別挑取含有pYES2空載體和pYES2-IbTPS的單菌落接種于10 mL SD-URA液體培養基中,30℃振蕩培養24 h,測定A600nm值,將菌液轉接至含有半乳糖(20 g/L)的誘導培養基中,使初始A600nm值均為0.4,30℃誘導表達30 h,將最終菌液A600nm值調為一致。取1 mL誘導的酵母菌液,離心后去上清液,菌體分別懸浮于1 mL的5 mol/L NaCl、4 mol/L山梨醇和PBS溶液(pH 7.4),置于4℃脅迫處理24 h,稀釋1 000倍,取50 μL涂布于不含半乳糖的SD-URA固體平板上,放至30℃培養箱中培養,通過平板計數計算釀酒酵母細胞的生存率。
1.2.7 數字表達譜分析 利用Illumina Pipeline軟件獲取甘薯的嫩葉、成熟葉片、莖、須根、初始塊根、膨大塊根和收獲塊根等7個不同組織原始的21 bp的DGE標簽,Bowtie定位有關IbTPS7a基因完整CDS的clean tag(僅允許一個堿基錯配)。使用edgeR package程序對RNA的組成和不同的測序庫規模進行誤差評估,對獲得的基因的表達量進行標準化,從而獲得準確的IbTPS7a基因的表達量[13]。
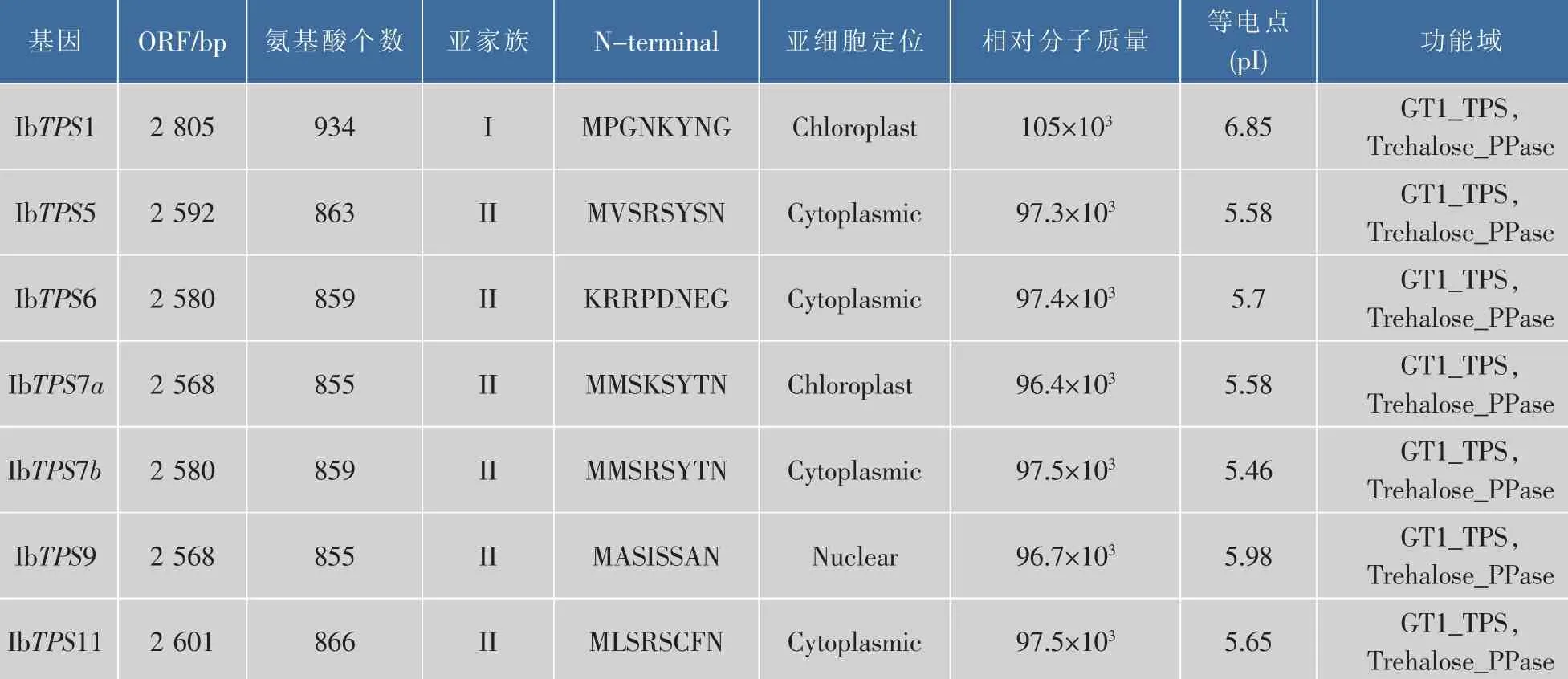
表1 甘薯的TPS基因家族Table 1 TPS family genes in Ipomoea batatas
2 結 果
2.1 甘薯TPS基因家族挖掘及分析
為了挖掘甘薯的TPS基因家族,作者根據擬南芥和水稻的TPS家族核酸及蛋白序列對甘薯轉錄組數據庫分別進行BlastN和BlastP,將比對得到的contig序列翻譯得到的TPS蛋白進行序列和功能單元分析,共篩選得到7個全長IbTPS基因(如表1所示),其中有兩個IbTPS蛋白序列經比對發現其同源性達到75%,且這兩個IbTPS蛋白與其它物種的TPS7蛋白序列同源性最高,因此分別被命名為IbTPS7a和IbTPS7b。生物信息學分析發現甘薯TPS蛋白都具有TPS蛋白所含有的N端GT1_TPS保守區域及C端Trehalose_PPase保守區域。信號肽分析結果表明7個甘薯TPS蛋白都沒有信號肽,亞細胞定位分析發現IbTPS1和IbTPS7a最有可能位于葉綠體 中 ,IbTPS5、IbTPS6、IbTPS7b 和 IbTPS11 可能位于細胞質中,而IbTPS9可能位于細胞核中,這表明甘薯不同的IbTPS蛋白在胞內分布區域及功能可能有所不同。利用已知的擬南芥11個TPS蛋白與甘薯7個TPS蛋白進行進化樹分析,結果發現與擬南芥TPS蛋白家族類似,甘薯的TPS蛋白也可以分為兩個亞家族,其中IbTPS1屬于I類家族,其余IbTPS蛋白屬于II類家族(如圖1所示)。
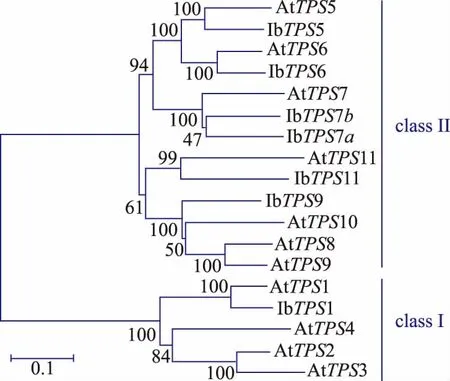
圖1 甘薯和擬南芥中TPS蛋白家族進化樹分析Fig.1 Phylogeneticanalysisofthe TPS family in Arabidopsis and Ipomoea batatas
2.2 IbTPS7a基因克隆及表達載體構建
以提取得到的徐薯18總RNA為模板,使用通用引物oligo(dT)15進行反轉錄得到cDNA,再以cDNA為模板使用TPS-32sF和TPS-32sR進行PCR,以 pET32a(+)質粒為模板使用 PET_TPS-F和PET_TPS-R進行擴增,電泳檢測在約2 600 bp左右和5 900 bp左右各有一明亮條帶(圖2),與之前預期的擴增的IbTPS7a基因大小和pET32a(+)質粒大小一致;使用SLIC法構建pET32-TPS質粒,以測序正確的pET32-TPS質粒為模板,使用TPS-yF和TPS-yR進行PCR擴增,在2 600 bp左右有一明亮條帶,將PCR片段與pYES2載體分別雙酶切后連接轉化,得到pYES2-IbTPS質粒。
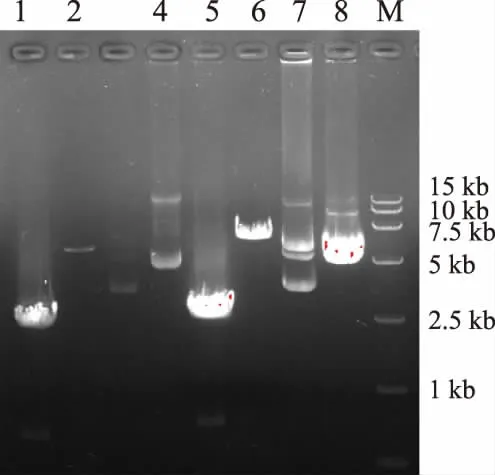
圖2 IbTPS7a基因PCR產物及載體構建Fig.2 Agarose electrophoresis analysis and expression vector construction of IbTPS7a
2.3 IbTPS7a生物信息學分析
生物信息學分析結果顯示,IbTPS7a基因長度為2 568 bp,其翻譯的蛋白長度為855 aa,IbTPS7a蛋白分子式為C4310H6710N1158O1277S40,蛋白相對分子質量為 94.46×103,理論等電點 5.58,脂肪系數為87.22,親水性值為-0.230,因此IbTPS7a是一個親水性蛋白;信號肽預測分析發現IbTPS7a沒有明顯的信號肽,可見IbTPS7a是一個胞內蛋白,主要在細胞內發生作用,跨膜結構分析發現IbTPS7a具有兩個明顯的由外到內跨膜結構區域:474-491和790-812;通過亞細胞定位軟件Wolf psort對IbTPS7a蛋白在甘薯細胞內的位置進行了預測,結果發現K最近鄰(k-Nearest Neighbor,KNN)值為 14,其中 7 個位于葉綠體中,5個位于細胞核中,2個位于液泡中,即IbTPS7a定位可能性為:葉綠體>細胞核>液泡。
2.4 IbTPS7a蛋白進化樹分析
使用DNAMAN軟件將10種其它植物的TPS蛋白與甘薯IbTPS7a蛋白進行序列比對,發現它們同源性都在82%以上,其中IbTPS7a與Nicotiana sylvestris和Solanum tuberosum蛋白同源性最高為86.7%,與Eucalyptus grandis的TPS7蛋白同源性最低為82%(如圖3所示)。將這11條蛋白序列首先進行MEGA 5.0軟件進行Clustal W比對,然后將比對后的蛋白序列采用Neighbor-joining法構建系統進化樹,Bootstrap自檢1 000次,甘薯IbTPS7a蛋白與Nicotiana sylvestris和Solanum tuberosum的TPS7蛋白親緣關系最近,該結果與同源比對結果一致。

圖3 IbTPS7a蛋白進化樹分析Fig.3 Phylogenetic tree of IbTPS7a protein
2.5 含IbTPS7a基因的大腸桿菌BL21重組子抗逆境脅迫分析
為了驗證IbTPS7a基因在原核細胞中的功能,將 pET32-IbTPS質粒轉化 E.coli BL21(DE3)并使用IPTG進行誘導表達,分別使用0.6、0.8 mol/L NaCl、質量分數20%PEG6000和30%PEG6000對IbTPS7a基因過表達菌株進行高鹽及干旱模擬脅迫,并對生長曲線進行測定。如圖4顯示,在無脅迫的LB培養基中含有空載體的對照菌株和轉IbTPS7a基因菌株的生長曲線幾乎沒有差異(圖4);在0.6 mol/L NaCl脅迫處理下,轉IbTPS7a基因菌株(0 h)相比對照菌株(4 h)更早進入指數生長期,但是兩者進入穩定期的時間差異不大,轉基因菌株生長效果較好于對照菌株;在0.8 mol/L NaCl脅迫處理下,兩種菌株的生長都受到抑制,但轉基因菌株的生長明顯優于對照菌株;在20%PEG6000脅迫處理下,兩種菌株的生長效果差異并不明顯;在30%PEG6000脅迫處理下,對照菌株生長困難,轉基因菌株明顯優于對照菌株,但是相比20%PEG6000脅迫轉基因菌株進入指數生長期的時間也明顯延長。
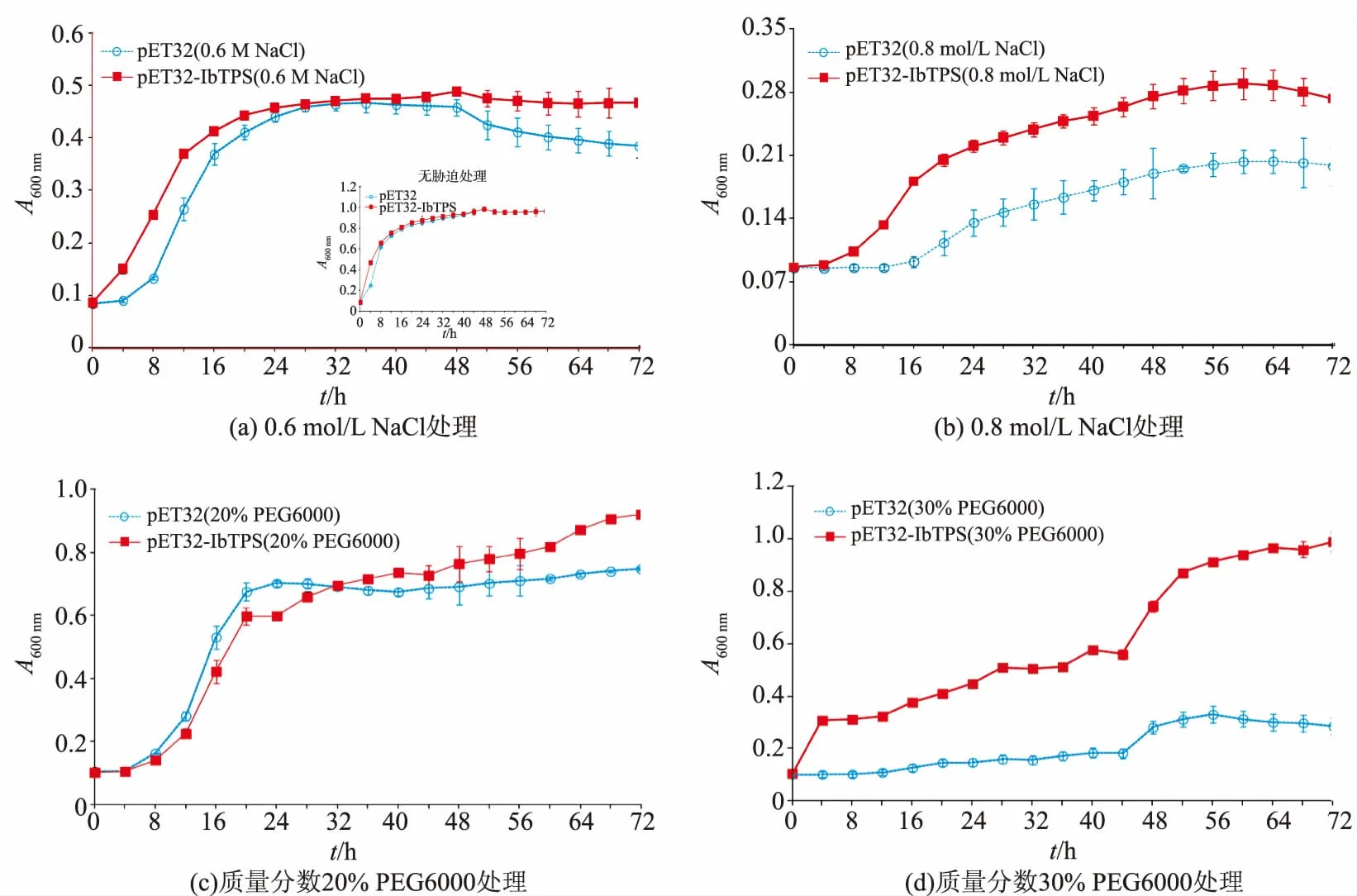
圖4 重組大腸桿菌BL21/IbTPS7a在鹽脅迫及干旱脅迫培養基中的生長曲線Fig.4 Growth curves of IbTPS7a in E.coli BL21 under salt and drought stress
2.6 轉基因酵母抗逆境脅迫分析
如圖5所示,在無脅迫壓力下,含有pYES2空載體的酵母菌株和轉IbTPS7a的菌株涂平板后的菌落數基本一致,這表明IbTPS7a基因的過表達對釀酒酵母的正常生長沒有明顯的影響;兩種菌株分別在5 mol/L NaCl和4 mol/L山梨醇進行脅迫處理后,轉IbTPS7a基因的釀酒酵母菌株的存活率明顯高于對照菌株,因此實驗結果表明IbTPS7a基因的過表達可以有效提升釀酒酵母的生存率,降低外界環境改變對細胞的傷害。
2.7 IbTPS7a基因數字表達譜分析
利用數字表達譜對甘薯IbTPS7a基因的組織特異性表達進行分析,結果(如圖6所示)發現,甘薯在多個組織中的表達量相對較低,其中在莖(ST)中的表達量最高為15.35 TPM,其次是嫩葉 (YL)為12.23 TPM,在根部組織的表達量都比較低,其中在須根(FR)中的表達量最低為0.84 TPM。
3 討論
TPS基因可通過多種信號傳導途徑參與植物的代謝、生長發育及抗逆作用,不同物種的TPS基因對脅迫條件的反應機制存在差異,TPS基因在不同的組織中的表達量也不盡相同[14-15]。甘薯是一種重要的糧食作物,其栽培品種是六倍體,基因組龐大且結構復雜,由于甘薯基因組測序尚未完成,現在對于甘薯遺傳資源的挖掘多通過轉錄組數據來實現[16-17]。在已完成基因組測序的高等植物中都發現有TPS基因家族,而目前尚沒有對甘薯TPS基因家族研究的報道。

圖5 重組釀酒酵母菌在鹽脅迫及干旱脅迫條件下的生長Fig.5 Growth of IbTPS7a in yeast under salt and drought stresses

圖6 IbTPS7a基因在甘薯不同組織樣品中的表達量Fig.6 Expression level of IbTPS7a in different tissues of Ipomoea batatas by digital gene expression profiling(TPM means the transcript per million)
4 結語
作者使用生物信息學方法對甘薯轉錄組數據庫進行比對分析,首次鑒定得到7個具有完整ORF結構的甘薯TPS基因家族成員,其中有兩個TPS基因經鑒定為TPS7基因的兩個亞型即IbTPS7a和IbTPS7b,本研究對于TPS基因家族的挖掘基于轉錄組數據,沒有找到全部的甘薯TPS基因,顯示了數據不足的缺憾,但是也為未來甘薯TPS基因的研究提供了重要的數據。將甘薯的TPS基因家族與其擬南芥的TPS基因進行了進化樹分析,結果發現與擬南芥類似,甘薯TPS基因家族也可以被分為兩個亞家族,其中IbTPS1歸屬于class I,其余的IbTPS基因屬于class II。擬南芥的TPS1被證明具有TPS活力,但class II的TPS尚未被證明具有活力[18],為了研究甘薯class II中TPS基因的功能,作者利用RT-PCR方法成功克隆得到class II的IbTPS7a基因,并對其在大腸桿菌和釀酒酵母中的抗逆作用進行的研究,結果發現轉IbTPS7a基因后誘導過表達的大腸桿菌菌株相比對照菌株在高鹽和干旱脅迫下生長能力明顯提升,在釀酒酵母中過表達IbTPS7a基因也可以有效增強真核細胞在高鹽和干旱脅迫下生存率也顯著提高,這說明過表達甘薯IbTPS7a有助于提升細胞的耐鹽和耐干旱作用,增強細胞適應外界環境變化及抵御脅迫的能力。細菌中海藻糖合成除了TreS和TreY/Z途徑外還可以通過TPP/TPS途徑,酵母中也具有TPP/TPS途徑,研究發現外源TPS基因的導入可以很好的修復相關缺陷菌株的海藻糖合成能力,因此甘薯TPS基因的過表達可能有助于促進大腸桿菌和酵母胞內海藻糖的合成,從而保護細胞免受環境脅迫造成的傷害[19-20]。大豆和擬南芥的TPS基因在根和根瘤等組織中的表達量較高[21],而作者發現IbTPS7a基因在莖和嫩葉中的表達量較高,但是在多個根部組織中的表達量較低,這可能是因為甘薯IbTPS7a基因與其它植物中的TPS基因在功能上有所不同。由于海藻糖對生物體優良的保護性能,其在食品、保健、醫療等行業應用越來越普遍[22],因此本研究不僅為進一步研究甘薯TPS基因家族及IbTPS7a的功能打下了基礎,也為拓展甘薯海藻糖合成酶在未來的應用提供了理論依據。
