南海氮循環幾個重要過程研究的新進展
2018-12-06 03:58:30楊進宇高樹基
廈門大學學報(自然科學版) 2018年6期
楊進宇,高樹基
(廈門大學 海洋與地球學院,近海海洋環境科學國家重點實驗室,福建 廈門 361102)
對于大多數海洋生態系統而言,氮元素是海洋初級生產力和輸出生產力的主要限制因子之一,其源匯格局對于調控海洋生物泵效率和大氣CO2濃度有重要意義[1].另外,氮循環過程中會釋放溫室氣體N2O,其溫室效應約為CO2的300倍,定量估算其釋放通量對于評估大氣溫室效應和全球氣候變化至關重要[2].工業革命(18世紀60年代)以來,人為排放的活性氮日益增加,其經河流和大氣傳輸進入海洋的通量已與海洋中生物固氮的通量相近[3-4].外源氮的輸入可能干擾海洋中不同氮組分的相互轉化,改變海洋氮庫的動態平衡,影響氮與其他生源要素循環的相互耦合關系,可能導致海洋生態系統的劇變,該現象在邊緣海將尤為顯著,從而引起關注[5].邊緣海環境因子多變,其內部氮循環過程,尤其是相關定量和作用機制研究,還存在諸多不確定性.此外,需要進一步探索和評估邊緣海氮循環過程對人為活動和氣候變化的響應及反饋.
南海是西北太平洋最大的邊緣海,具有典型的熱帶-亞熱帶氣候特征,其表層環流和生物地球化學過程主要受東亞季風的影響[6].南海被人口密集的亞洲大陸和島嶼包圍,形成半封閉狀態,周邊河流攜帶大量陸源物質匯入[7];另一方面,南海海盆區屬于大洋主控型邊緣海,經呂宋海峽這唯一的深水通道與開闊的西北太平洋進行水體和營養物質的交換[8-9].此外,南海及其周邊海域也是全球受到人為活動影響最為顯著的區域之一[4].諸多過程相互影響使得南海的氮收支和內部循環過程相對復雜;同時,也使南海成為探究邊緣海氮循環過程非常好的試驗場[10].隨著同位素示蹤技術的發展和成熟,其在南海氮循環研究中被廣泛應用.本文中針對南海外源氮輸入的變化,上層水體中氨氮動力學過程,以及顆粒態氮動力學過程的相關研究進行總結和歸納,重點展示南海氮源、氮的內部循環和氮匯等過程的新進展.
1 南海外源氮輸入的變化
1.1 南海外源氮輸入通量
外源氮是指從海洋外部進入海洋內部的新生氮,外源氮輸入能夠有效地改變海-氣界面CO2通量穩態平衡[4].對于邊緣海來說,河流輸入、固氮作用和大氣氮沉降是最為重要的外源氮輸入途徑(表1).南海周邊河流輸入無機氮通量約為64.0 Gmol/a, 其中珠江和湄公河總貢獻量高達62%[11];而河流輸入的有機氮通量僅為33 Gmol/a[18].河流攜帶的大量營養物質具有較高的氮磷比(摩爾比高達80∶1)[18],可能緩解或改變南海氮限制的狀態.然而,Voss等[12]的研究顯示河流的影響會刺激固氮作用的發生,因此河流攜帶的其他生源要素(如鐵、鎳等)可能發揮重要作用.更多的研究顯示河流僅能影響近岸陸架區的生物活動和有機物埋藏[12,19],其對南海開闊海區的影響則相對有限.南海海盆區(面積約為1.4×106km2)全年大部分時間處于寡營養鹽狀態[20],其高溫、層化等環境條件有利于固氮生物生長[21].采用不同方法估算的南海總固氮速率存在較大的時空變異,約為3~30 Gmol/a,其中單細胞固氮藻貢獻約65%的固氮速率[12-15].然而,南海固氮藻的豐度和生長速率均低于鄰近的西菲律賓海[13,15].Wu等[22]推測南海表層水體生物可利用鐵的含量較低,從而限制了南海固氮生物的生長.此外,南海較高的大氣氮沉降通量也可能是影響固氮生物生長的因素之一.Yang等[16]在南海北部東沙島的觀測顯示大氣沉降提供的總活性氮為55~70 mmol/(m2·a)(77~98 Gmol/a),這與Kim等[17]根據南海周邊陸地上觀測的推導結果以及Duce等[4]利用模型估算的入海氮沉降通量相似,但遠高于早期的觀測結果(7~9 mmol/(m2·a))[18].上述結果表明南海的大氣氮沉降通量明顯高于固氮作用固定的氮通量,貢獻約20%的新生產力,是海盆區最主要的外源氮輸入途徑.南海大氣沉降中硝酸鹽是主要成分,其來源存在明顯的季節變化,與季風有密切關系,表現為:在冷季以人為活動來源為主,而在暖季可能來源于閃電合成和土壤釋放等天然過程以及生物體焚燒[16,23].目前對南海大氣中還原態氮(銨鹽和有機氮)的主要來源了解較少.有限的研究顯示在北大西洋大氣中還原態氮主要源自海洋,說明大氣中人為活動來源的氮貢獻可能存在高估[24-25].北大西洋大氣中人為活動來源的氮主要源自北美大陸的排放,其主要成分是氮氧化物(NOx)[26];而由東亞大陸產生的人為排放氮組分中氮氫化物(NHy)的貢獻可能超過50%[27].因此,南海甚至整個北太平洋的大氣氮沉降中還原態氮的來源可能與北大西洋存在差異,值得深入研究.未來在南海及周邊海域開展大氣中還原態氮的穩定氮同位素和有機組成研究將有助于準確評估外源氮輸入的貢獻以及人為活動的影響.
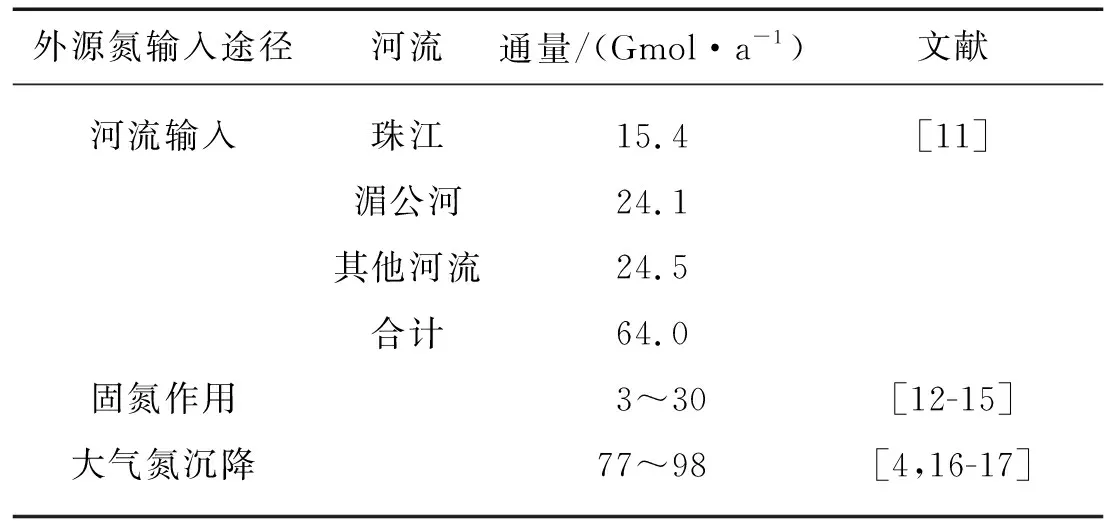
表1 南海外源氮輸入通量
1.2 人為活動對南海外源氮輸入的影響
外源氮輸入可能顯著影響南海內部的生物地球化學過程.總體上,南海是一個氮限制的生態系統[28-29],外源氮輸入的長期變化對于調控南海新生產力有重要意義.人為活動對河流近岸區的影響尤為明顯.以珠江口區域為例,近20多年來營養鹽濃度明顯上升,尤其是氮鹽((1.4±0.3) μmol/(kg·a)),可能加劇了近岸區域的富營養化和缺氧狀態[30-31],由此可能增強近岸水體的反硝化過程和溫室氣體(N2O)的釋放.此外,Ren等[32]在南海根據珊瑚體內穩定氮同位素記錄成功反演出人為活動經大氣沉降對海洋氮儲庫的影響,結果發現過去40年珊瑚礁體內穩定氮同位素組成(δ15N)的減少與大氣氮沉降(主要是燃煤引發)的增加密切相關.現階段在南海由大氣氮沉降提供的外源氮能夠增加約460 mmol/(m2·a)的CO2吸收量(以光合作用中n(C)∶n(N)=6.6計算),與南海向大氣釋放的CO2通量[33]處于同一量級.這說明在工業革命之前(人為活動影響較弱),南海釋放的CO2通量可能是現在的2倍.類似地,模型研究顯示未來人為活動的影響還將持續增加[4],南海作為CO2弱源的現狀是否會改變,值得關注[16].沉積物中有孔蟲體內的δ15N記錄顯示冰期與間冰期的固氮作用存在明顯差異,全新世較強的固氮作用主要由水溫變暖和層化加強所致[14].在全球變暖和海洋酸化的大環境下,固氮速率可能呈現下降的趨勢,在南海這種鐵限制的海域可能更為顯著[34].然而,南海大氣氮沉降可能進一步對固氮生物生長產生負反饋,這需要在今后的研究中加以驗證.
2 南海上層水體的氨氮動力學過程
上層水體營養鹽主要來源于有機物礦化產生的氨氮,以及外源氮輸入和次表層輸入的新生氮(硝酸鹽為主).傳統上認為浮游植物生長時吸收氨氮為再生生產力,而利用硝酸鹽的生長定義為新生產力.新生產力的大小決定了有機物輸出的效率.然而,氨氮氧化產生硝酸鹽進而被浮游植物利用,這一過程可能造成新生產力的高估[35];同時,還將直接影響上層海洋的氮儲庫形態,進而影響浮游植物群落結構.因此,準確定量地研究上層水體中氨氮的動力學過程非常重要.
以往的研究顯示表層水體的硝化作用會受到光抑制[36].然而,Wan等[37]在南海的研究發現:真光層內部氨消耗途徑與硝酸鹽濃度有關,并指出通過影響不同浮游植物種群對氨氮的親和力,可以改變不同營養鹽濃度下浮游植物和硝化菌對氨氮的競爭力.該研究成功地揭示了硝化作用被光抑制的假說與真光層內部活躍的硝化過程發生的矛盾,能夠廣泛適用于其他寡營養鹽海域(圖1).此外,該研究也更深入地闡釋和預測了硝化作用和新生產力的緊密空間耦合關系,有助于定量評估真光層內部的硝化作用對新生產力估算的影響.
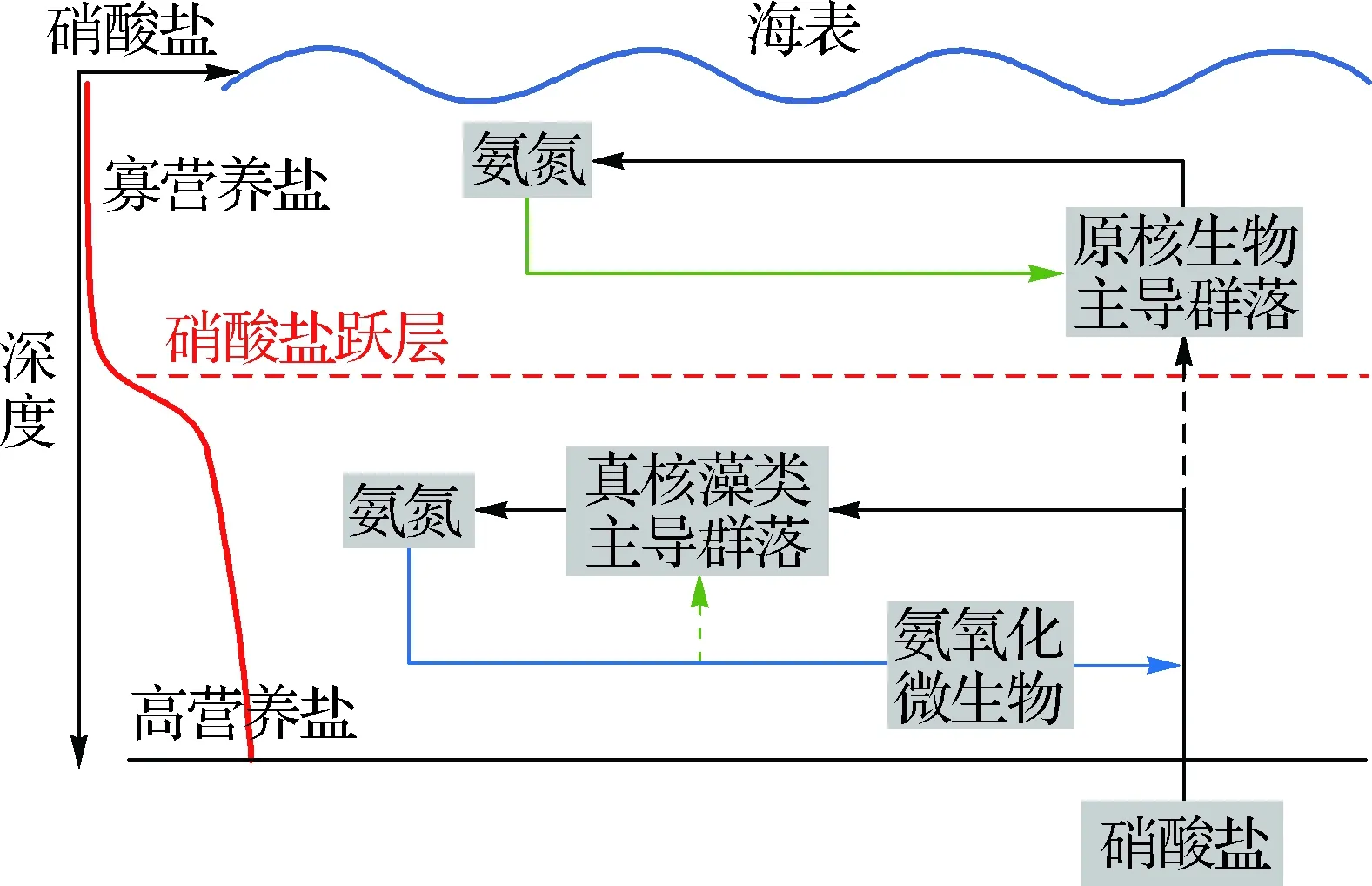
綠色箭頭表示氨氮的吸收過程,藍色箭頭表示硝化作用.
此外,南海上層水體的氨氮動力學過程還受到開闊大洋的影響.Xu等[38]揭示了在南海北部氨氧化速率的分布,發現氨氮濃度及其氧化速率與黑潮入侵有關:黑潮攜帶高含量的溶解有機物,在南海內部被分解從而提高了氨氧化速率,并為生物生長提供了額外的營養物質.該現象可能普遍存在于水平傳輸混合強烈的區域,如受黑潮和灣流等西邊界流影響的東海和墨西哥灣區域,未來需要開展更多的研究來證實這一推測.
南海上層水體中N2O處于過飽和狀態,是N2O的源(向大氣釋放19.3 Mmol/d),而與之相鄰的西菲律賓卻是N2O的匯[39].上層水體的硝化作用可能是南海上層水體中N2O的主要產生過程.模型預測上層海洋層化將進一步加劇,從而限制下層營養鹽的供給,未來以微微型原核生物為優勢種的寡營養鹽海域可能進一步擴大[40].這可能導致上層水體氨氮吸收速率增大,進而減弱硝化作用.然而,營養鹽躍層深度的變化又會改變硝化菌的活躍空間.因此,南海N2O的源匯格局變化值得關注.
3 南海顆粒態氮動力學過程
3.1 上層水體顆粒態氮同位素組成
邊緣海的顆粒有機物(POM)除了來源于海洋生物的現場生產,陸源物質和大氣沉降等也可能具有顯著貢獻.POM的穩定同位素信號能夠用來指示不同氮源,進而了解POM在水體中的動力學過程,以及反演現代和過去海洋的氮循環過程[41].通常情況下,邊緣海由于具有較高的生產力和顆粒沉降速率,顆粒物的同位素信號能夠較好地保存在水體和沉積物中[42-43].然而,POM從生成到埋藏經歷諸多復雜過程,穩定氮同位素信號對充分了解邊緣海中POM沉降過程中的控制因素至關重要.Yang等[44]在南海北部海盆區的研究發現:在真光層底部收集的沉降顆粒物與次表層硝酸鹽具有相似的δ15N(均為4×10-3~5×10-3),表明次表層硝酸鹽是提供顆粒物輸出最主要的新生氮源;進一步比對發現上層100 m水體中不同種類POM(懸浮和沉降顆粒物,以及不同粒徑的浮游動物)具有高度相似的δ15N,暗示上層水體中氮元素在不同顆粒組分間快速運轉[44].這與在其他寡營養鹽海域的研究結果有所不同,如Montoya等[45]在北大西洋發現懸浮顆粒物的穩定氮同位素組成(δ15NPN)明顯低于沉降顆粒物的穩定氮同位素組成(δ15NSP),且浮游動物的穩定氮同位素組成(δ15Nzoo)隨粒徑增加而增大.盡管造成上述差異的控制機制尚不清楚,但推測在南海溶解有機物氮可能有效地參與了上層水體氮動力學過程[46].Kao等[14]在南海北部海盆區發現上層100 m水體中懸浮顆粒物的氮含量和δ15NPN(2×10-3~6×10-3)存在明顯的季節變化,春、秋兩季較低的δ15NPN對應于較低的顆粒物向下輸出效率,暗示固氮作用可能是主導因素.因為大多數固氮藻(南海以固氮變形菌為主[47])的沉降速率較慢,所以具有較輕同位素信號(δ15NPN較低)的顆粒物在上層水體中的停留時間較長.固氮作用信號是否能夠有效地向下輸出,以及以何種形式輸出,需要進一步深入研究.相反地,在冬季或夏季臺風期間,較強的水體混合常引發中等規模的硅藻藻華[48],具有下層硝酸鹽同位素信號的顆粒物能夠快速有效地向下輸出,從而造成真光層內部顆粒有機氮的儲量反而較低[18].
3.2 顆粒態氮同位素信號的垂直分布
不同于其他邊緣海,南海的POM在向下輸出過程中似乎沒有完好地保存原有的同位素信號特征(圖2).研究發現南海深層水體中δ15NSP為2×10-3~3×10-3[44,49],低于真光層的δ15NSP和次表層硝酸鹽穩定氮同位素組成(δ15NNO3).根據沉積物捕獲器的數據顯示,南海在3 000 m以下收集到的顆粒氮通量可能高于上層水體,說明存在由陸架區側向輸入的POM的貢獻[44].如Huh等[50]報道臺灣島東南部高屏溪每年往南海陸架匯入36~49 Mt顆粒物,其中約85%經高屏峽谷輸送到南海深海海盆;Liu等[51]通過分析海盆區沉積物中黏土礦物發現南海北部的顆粒物主要來源于臺灣島河流.這些側向輸入海盆的POM往往具有較輕的氮同位素特征[44].另外,Jiao等[52]首次在南海深層水中發現生活在表層水體中的原綠球藻(Prochlorococcus),推測是南海內部常發生的內孤立波等物理過程導致POM從表層向深層輸出.這類單細胞原核生物具有較低的δ15N[53],因而這也是使得南海深層水中δ15NSP變低的可能機制.然而,至今還沒有針對南海區域不同種群微生物的穩定氮同位素研究,尚無法評估該機制的重要性.未來加強流式分析-同位素質譜、特征氨基酸組分及其同位素組成的分析應用將有助于理解水柱中沉降顆粒物氮同位素信號的傳輸和轉化過程.
深層水體中δ15NPN和δ15NSP表現出顯著差異.Saino等[54]發現南海深層水中δ15NPN增加至8×10-3左右,明顯高于δ15NSP.兩者的顯著差異表明較小的懸浮顆粒物不是完全由較大的沉降顆粒的物理分解或水解產生的,兩者可能有不同的來源.這與氨基酸組分[55]和蛋白組學[56]的分析結果相似,其結果顯示深層懸浮顆粒物的來源有別于沉降顆粒物和表層懸浮顆粒物.Gaye等[55]推測深海中細小顆粒物(如懸浮顆粒物)的穩定氮同位素是由于其停留時間較長,與溶解有機物交換所致.開展全水柱的溶解有機物以及有機組分的δ15N研究有望驗證上述假設.
Kienast[57]發現南海表層沉積物和巖芯中δ15N在上一個冰期-間冰期時相對穩定.然而,這一結果有悖于有孔蟲體內同位素變化的分析結果[14].南海水體中δ15NSP的顯著變化也表明在該區域利用其反演氮循環的歷史變化可能存在問題.冰期-間冰期期間海盆區陸源物質的來源和貢獻量存在較大變化[51],因此可能混淆沉積物中不同時期的氮同位素記錄[58].上述發現對過去在邊緣海重建氮循環過程的認識提出了新的挑戰[38].
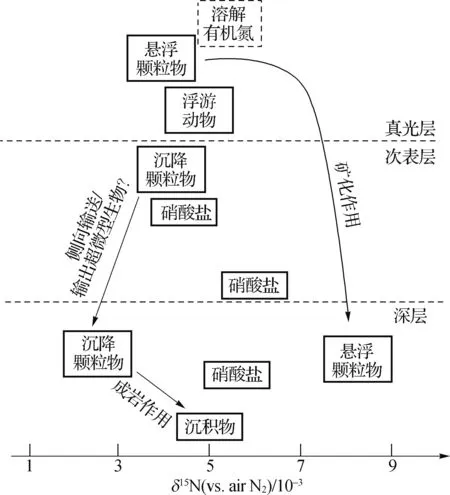
其中溶解有機物的δ15N為估計值.
4 展 望
南海氮循環研究起步較晚,盡管近年來已經取得了長足進步,但仍需在以下幾個方面加強研究:
1) 硝酸鹽同位素組成及其控制機制.硝酸鹽是海洋中最大的固定氮儲庫,系統地分析其穩定氮、氧同位素的組成及時空分布有助于了解南海硝酸鹽的動力學過程.北太平洋中層水最終進入南海[59],通過硝酸鹽同位素可指征其歸宿,這對反演南海和北太平洋氮循環在現代和歷史上的變化有重要意義.
2) POM和溶解有機物的有機組分及其穩定氮同位素.這有助于厘清溶解有機物降解的分餾效應,揭示溶解有機物在參與水體中不同氮組分的相互轉化中的作用,以及不同粒徑顆粒物的來源和相互關系.
3) 南海上層水體硝化作用的季節變化.南海上層水體的營養鹽水平和躍層及黑潮入侵南海的強度存在明顯的季節變化,因此需要加強不同季節硝化作用強度及其對南海浮游植物群落、生物泵效率的影響,以及對N2O產量的關系和貢獻等方面的研究.
4) 人工標記技術在氮循環過程研究中的應用.目前,利用不同氮組分的同位素人工標記技術能夠有效地定量不同氮循環過程的速率,有助于揭示不同氮循環過程與多環境因子的相互關系[60],該技術應該被廣泛應用于現場工作中.
5) 南海南部的氮循環過程.目前大多數研究還集中在南海北部,對南部的認識相當匱乏.南海南部是南海水團年齡最老的區域,應關注其與北部海域的對比研究.
6) 氣候變化與人為活動對氮循環過程的影響.氣候變化和人為活動使得海洋生物地球化學過程處在持續變化中,氮循環過程亦是如此.關注氮循環過程對外界環境的響應和反饋將是今后研究的重點.
猜你喜歡
軍事文摘(2023年10期)2023-06-09 09:15:06
空間科學學報(2021年4期)2021-08-30 08:31:18
環境保護與循環經濟(2017年2期)2017-09-26 11:52:22
環境科技(2016年1期)2016-11-08 12:17:48
河北書畫研究(2016年2期)2016-08-24 02:14:50
新農業(2016年18期)2016-08-16 03:28:27
化工進展(2015年3期)2015-11-11 09:18:15
浙江大學學報(工學版)(2015年1期)2015-03-01 01:17:28
環境與可持續發展(2013年6期)2013-03-11 16:21:48
環境與可持續發展(2013年3期)2013-03-11 16:21:18
