2013—2016年我院主要革蘭陰性菌對頭孢西丁的耐藥性分析
2018-12-20 02:36:02邱剛沈明張林祥朱海英趙穎斐宋美琳
浙江醫學 2018年23期
關鍵詞:耐藥
邱剛 沈明 張林祥 朱海英 趙穎斐 宋美琳
頭孢西丁是由美國默沙東公司研發的頭霉素類抗菌藥物。頭孢西丁鈉由于在β-內酰胺環7號位上引入α-甲氧基,使其對革蘭陰性菌產生的β-內酰胺酶所引起抗菌藥物失活起到了重要保護作用,其抗菌譜廣,對革蘭陽性、陰性需氧及厭氧致病菌均有抗菌活性。隨著抗菌藥物的大量應用,多重耐藥菌不斷出現,具有耐超廣譜β-內酰胺酶(ESBLs)特點的頭霉素類藥物重新被臨床所重視。筆者對我院2013—2016年頭孢西丁的臨床使用分布情況及臨床分離的革蘭陰性菌耐藥性進行分析,現將結果報道如下。
1 資料和方法
1.1 一般資料 從我院合理用藥軟件導出2013-2016年各科室住院患者抗菌藥物使用數據,包括抗菌藥物名稱、規格、消耗量、銷售金額、用藥頻度和同期收治患者住院人/天數等。菌株分離自2013—2016年臨床收集的各種標本,包括血液及骨髓、中段尿、痰液、腦脊液、分泌物、咽拭子等。
1.2 方法 采用世界衛生組織(WHO)推薦的限定日劑量(DDD)方法,DDD值依據《新編藥物學》第17版和《中國藥典》(2015年版)及藥品說明書取其主要適應證劑量確定。用藥頻度(DDDs)=某藥品的年消耗總量/相應的DDD值,其值越大,說明使用頻率越高,選擇傾向性越強。
1.3 藥敏試驗及培養基 按衛生部《全國臨床檢驗操作規程》進行,菌種鑒定及藥敏試驗采用ATB自動微生物鑒定及藥敏試驗分析系統,藥敏試驗采用K-B紙片瓊脂擴散法。ESBLs確定根據美國臨床和實驗室標準協會(CLSI)推薦的雙紙片協同試驗,使用頭孢他啶、頭孢他啶/克拉維酸和頭孢噻肟、頭孢噻肟/克拉維酸確證試驗確認。
2 結果
2.1 2013—2016年我院頭孢西丁使用分布情況統計2014年我院頭孢西丁用量較上一年突增58.73%,之后2年使用量持續下降。普外科使用量近3年下降明顯,婦產科用量總體呈上升趨勢,兩者占全院頭孢西丁總使用量的81.88%以上。見表1。

表1 2013—2016年該院頭孢西丁使用分布情況統計(DDDs)
2.2 2013—2016年普外科頭孢西丁使用情況統計2013—2016年普外科頭孢西丁使用量占科室抗菌藥物總使用量的7.97%~24.51%,2014年較2013年用量增加了52.58%,之后2年又連續下降,見表2。根據合理用藥軟件導出的數據查找原因,發現后2年第3代頭孢類藥物(加酶抑制劑)以及甲硝唑使用量相應明顯增加。
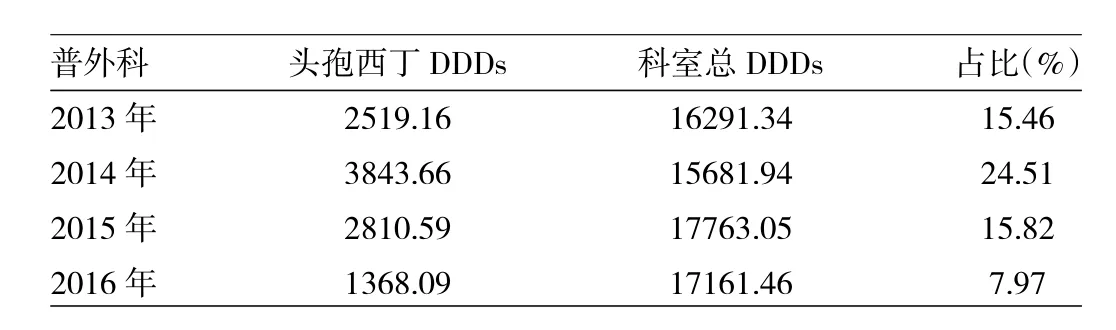
表2 2013—2016年普外科頭孢西丁使用情況統計
2.3 2013—2016年婦產科頭孢西丁使用情況統計2013—2016年婦產科頭孢西丁使用量占科室抗菌藥物總使用量的19.01%~38.44%。頭孢西丁使用量在科室抗菌藥物總使用量的占比逐年升高,說明臨床醫師在選用抗菌藥物時優先考慮選用頭孢西丁。見表3。
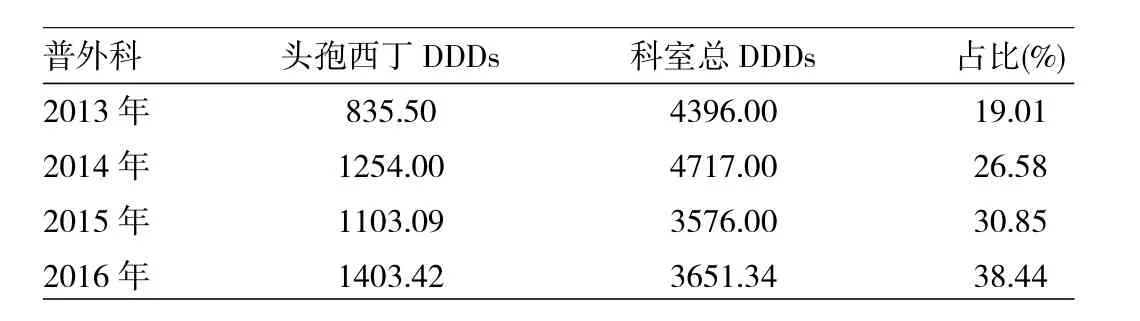
表3 2013—2016年婦產科頭孢西丁使用情況統計
2.4 2013—2016年分離的主要革蘭陰性菌對頭孢西丁的耐藥性統計 銅綠假單胞菌和鮑曼不動桿菌對頭孢西丁耐藥率較高(94.80%~98.51%),大腸埃希菌和肺炎克雷伯菌對頭孢西丁敏感性較高(2.29%~16.30%)。但2014年較2013年耐藥率及耐藥株數明顯上升。見表4。
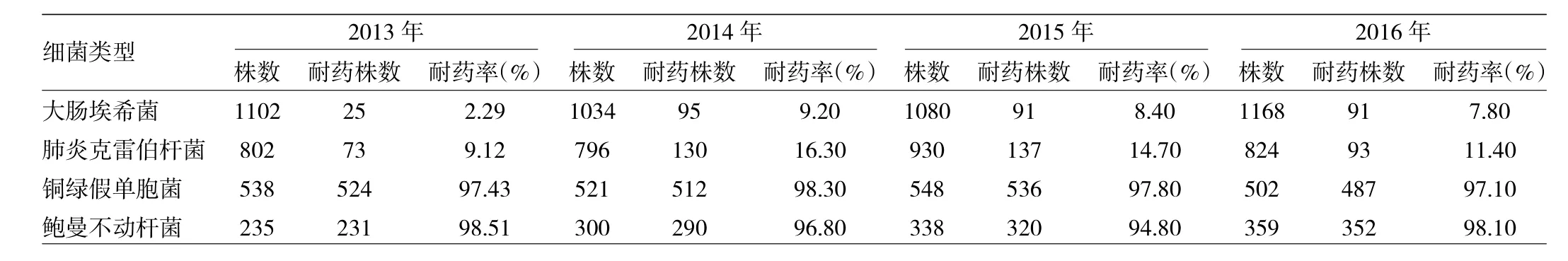
表4 2013—2016年分離的主要革蘭陰性菌對頭孢西丁的耐藥性統計
3 討論
本研究發現,近幾年我院普外科與婦產科頭孢西丁使用量占全院頭孢西丁總使用量的81.88%以上,這與頭孢西丁具有抗菌譜廣、對β-內酰胺酶穩定且兼有抗厭氧菌作用的特點以及科室、手術部位細菌分布有關。普外科2015年第3代頭孢類藥物頭孢地嗪和頭孢噻肟使用量相應的增加明顯以及2016年頭孢噻肟和頭孢哌酮舒巴坦用量上升明顯,說明臨床醫師已傾向于選用起點高的廣譜抗菌藥物。有文獻報道,第三代頭孢菌素的廣泛使用和濫用,是導致產ESBLs菌株增多的重要原因[1-3]。有調查發現,該院2013—2015年在普外科患者傷口分泌物中產ESBLs大腸埃希菌檢出率逐年上升(29.28%~34.97%)[4],《抗菌藥物臨床應用指導原則(2015年版)》指出,頭霉素類對大多數超廣譜β-內酰胺酶穩定,但其治療產ESBLs的細菌所致感染的療效未經證實,從而導致普外科近幾年將頭孢西丁作為次選藥物,這可能是頭孢西丁使用量近3年下降明顯的原因之一。
本研究發現,婦產科頭孢西丁使用量在科室抗菌藥物總使用量的占比逐年升高,說明婦產科臨床醫師在選用抗菌藥物時優先考慮選用頭孢西丁。根據《抗菌藥物臨床應用指導原則(2015年版)》頭孢西丁作為婦科子宮切除術圍術期預防用藥,可能是導致婦科使用量上升的原因之一,但在實際用藥醫囑中發現部分病歷存在頭孢西丁與甲硝唑等抗厭氧菌藥物合并使用,其中抗厭氧菌譜重疊。故在使用過程中應嚴格掌握抗菌藥物使用指征,避免無指征用藥。同時加強對頭孢西丁合理使用的監測與評估,從而減少不合理使用的情況。
本研究中銅綠假單胞菌對頭孢西丁的耐藥率在97%以上,因其本身具有AmpR基因,該基因低水平表達可以產生少量頭孢菌素(AmpC)-β內酰胺酶;當暴露于頭孢菌素類藥物時,AmpR基因激活,高產AmpC酶。AmpC酶可由染色體或質粒介導,產AmpC酶的細菌通常只對第四代頭孢菌素和碳青霉烯類藥物敏感[5]。鮑曼不動桿菌對頭孢西丁的耐藥率在94%以上,其對β-內酰胺酶耐藥機制較為復雜,涉及到產ESBLs、產金屬-β-內酰胺酶、產AmpC酶和產苯唑西林酶等[6]。曹小利等[7]對頭孢西丁不敏感肺炎克雷伯桿菌進行研究發現,對頭孢西丁不敏感肺炎克雷伯桿菌大多產ESBLs酶和肺炎克雷伯桿菌碳青霉烯酶(KPC酶),并伴有AmpC酶的流行。產AmpC酶菌,在持續使用頭孢西丁治療過程中,腸桿菌科細菌AmpC酶的耐藥性逐漸增高,頭孢西丁對腸桿菌科細菌AmpC酶感染的抑菌作用逐漸減弱,有必要針對腸桿菌科細菌AmpC酶感染的耐藥性進行深入研究和分析,為腸桿菌科細菌AmpC酶感染患者選擇合適的抗菌藥物治療[8-10]。因此,臨床應加強對頭孢西丁合理應用的管理,產生AmpC酶是細菌對頭孢西丁耐藥的一個重要原因,且產AmpC酶菌可作為一個潛在的傳播源,應加強對產AmpC酶株的檢測和監測,有助于臨床合理選用抗菌藥物。
綜上所述,我院分離的4種主要革蘭陰性菌中銅綠假單胞菌和鮑曼不動桿菌對頭孢西丁高度耐藥,而大腸埃希菌和肺炎克雷伯桿菌目前對頭孢西丁仍然較為敏感。對頭孢西丁耐藥可能提示細菌高產AmpC酶,或同時產ESBLs和AmpC酶[11]。雖然AmpC酶被認為是革蘭陰性菌尤其是腸桿菌科細菌產生耐藥的重要機制,但隨著更深入的研究發現膜通透性降低、泵外排作用和青霉素結合蛋白的改變等機制在細菌產生耐藥過程中也起著至關重要的作用,同時也正是存在各種機制的協同作用導致了耐藥現象更為嚴重[12]。由于本次研究未能對耐頭孢西丁菌株作進一步產AmpC酶菌的檢測及耐藥性研究,以致于未能得到相關數據來證實,在今后的研究中將對此作進一步的研究。
猜你喜歡
保健醫苑(2022年5期)2022-06-10 07:46:38
現代臨床醫學(2022年3期)2022-06-06 07:59:40
昆明醫科大學學報(2022年1期)2022-02-28 07:43:40
天津醫科大學學報(2021年3期)2021-07-21 09:04:02
科學大眾(2020年12期)2020-08-13 03:22:22
云南醫藥(2019年3期)2019-07-25 07:25:10
現代檢驗醫學雜志(2016年1期)2016-11-12 13:19:40
國外醫藥(抗生素分冊)(2016年6期)2016-07-10 11:34:45
中國衛生標準管理(2015年14期)2016-01-15 02:58:37
中國當代醫藥(2015年17期)2015-03-01 02:03:58
