組蛋白H3K4甲基轉移酶KMT2D研究進展
2018-12-26 06:30:46閻晗于明航尹潔王璽
天津醫科大學學報 2018年6期
關鍵詞:小鼠
閻晗,于明航 綜 述,尹潔,王璽 審 校
(天津醫科大學細胞生物學系,天津 300070)
表觀遺傳學在調控基因表達上扮演重要角色。組蛋白H3K4的甲基化是細胞修飾啟動子和增強子的1種方法。組蛋白H3K4的三甲基化(H3K4me3)主要聚集在轉錄活躍的啟動子區域,而一/二甲基化(H3K4me1,H3K4me2)則主要集中在增強子區域[1]。酵母菌中,Set1復合物中的催化亞基SET1,具有組蛋白H3K4的甲基轉移酶活性,參與組蛋白H3K4的 me1,me2,me3 的修飾[2-3]。在果蠅中,dSet1、Trx和Trr是已知的3種Set1樣組蛋白H3K4甲基轉移酶。哺乳動物體內存在6種Set1樣H3K4甲基轉移酶,分別是 KMT2A(MLL1)、KMT2B(MLL2)、KMT2C(MLL3)、KMT2D(MLL4、ALR、MLL2)、KMT2F(SET1A)和KMT2G(SET1B)。KMT2D對于基因調控起到重要作用,其突變導致多種發育疾病和腫瘤的發生。本文對哺乳動物中最主要的H3K4單甲基化轉移酶-KMT2D進行探討,以期為探究疾病治療的新方法提供思路。
1KMT2D蛋白
如圖1所示,KMT2D蛋白的結構包括N-末端的兩組植物同源結構域(PHD)(每組包含3個PHD),C-末端的SET結構域負責組蛋白H3K4的甲基化[3]。體外實驗表明,第2組PHD(PHD4-6)識別核小體的H4尾巴,對于KMT2D-催化組蛋白甲基化起決定作用[4]。SET結構域的突變和缺失導致KMT2D蛋白的不穩定,表明SET區域不僅具有酶催化活性,并且對于維持蛋白穩定性至關重要[5-6]。在SET結構域上游是一個PHD區域和富含苯丙氨酸和絡氨酸(FY-rich)N/C-末端(FYRN 和 FYRC)區域。此外,還包含一個高速泳動族非組蛋白(HMG-1)和九個核受體相互作用基序(LXXLLs)[7]。體外實驗表明,氨基酸Y5426和Y5512對于人KMT2D的酶活性起決定性作用[6]。人胚胎干細胞(ESCs)中KMT2D的Y5426突變后,KMT2D會失去酶活性,但并不影響KMT2D蛋白的穩定性[5]。

圖1 人類組蛋白H3K4甲基轉移酶KMT2D蛋白
2KYM2D蛋白復合物
KMT2D是首次在Hela細胞核提取物一個蛋白復合體中分離純化出來的。對Pax反式激活結構域作用蛋白(PTIP,一種與組蛋白H3K4甲基轉移酶類相關的蛋白)相互作用蛋白的親和純化研究表明,PTIP除了與DNA損傷應答有關的蛋白質相互作用外,還與 ASH2L、RbBP5、WDR5、DPY30、NCOA6、UTX(KDM6A)、PA1、KMT2C和KMT2D有關聯[8](圖2)。純化的KMT2C和KMT2D表現出強H3K4甲基轉移酶活性[9]。WDR5、RbBP5、ASH2L 和 DPY30 形成4個亞基的子復合物WRAD,此復合物對于所有哺乳動物中Set樣組蛋白甲基轉移酶復合物的酶活性起關鍵作用[10]。WDR5直接與KMT2C和KMT2D的C-末端FYRN/FYRC區域結合[8]。PTIP和PA1是KMT2C和KMT2D復合物中獨特的亞基,除此之外還包含一個特殊的亞基UTX,它介導組蛋白H3K27的去甲基化[11]。缺失KMT2D將導致細胞中KMT2D復合物的解體和UTX的不穩定。
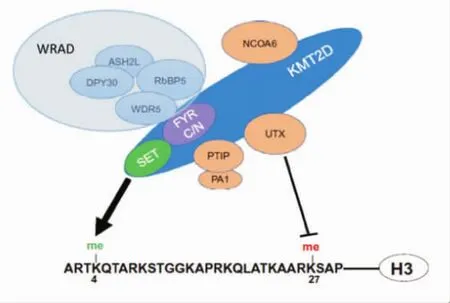
圖2 KMT2D蛋白復合物
3 KMT2D主要催化組蛋白H3K4一甲基化
增強子是順式作用元件,常與細胞型特異轉錄因子(TFs)結合,在真核細胞中對于特異基因的表達至關重要[12]。研究表明,KMT2D主要是通過其酶活性對細胞增強子區域進行H3K4—甲基化修飾,與KMT2C有功能冗余[13]。Lee等[13]運用ChIP-Seq研究,表明KMT2D在不同的細胞類型和分化水平,選擇性的結合在特異增強子區域。在細胞分化期間,轉錄因子(TFs)募集KMT2D與特異增強子結合。在未分化的KMT2C敲除細胞中,KMT2D基因缺失引起H3K4me1/2和H3K27乙酰化水平降低,轉錄輔助因子和RNA聚合酶II結合增強子的能力減弱,導致基因表達和細胞分化受阻。此外,KMT2D還識別超級增強子。在細胞分化過程中KMT2C和KMT2D對于超級增強子的形成是必須的[14]。
KMT2C和KMT2D對于H3K27乙酰轉移酶CREB-結合蛋白(CBP)和/或p300與增強子區域的結合、激活增強子、增強子-啟動子成環、活化啟動子,這一系列生物學過程是必須的[14-15]。由于乙酰化和甲基化往往是互相排斥的,可以推論,KMT2C和KMT2D復合物中的UTX亞基催化組蛋白脫甲基形成一個H3K27位點后,CBP和/或 p300對H3K27位點乙酰化(H3K27ac)。但是,研究表明UTX的H3K27脫甲基酶活性對于增強子活化和細胞特異基因表達是非必須的,其功能主要是與KMT2C和KMT2D形成復合物后作用于增強子[16]。
H3K4me1、H3K27ac是活化的增強子的標志[1],基于此論斷,提出KMT2D與增強子作用的三步式模型,如圖3所示,過氧化物酶體增殖物活化受體-γ(Ppar-γ)的增強子活化過程。(1)轉錄因子,如C/EBPβ,與增強子區域結合。(2)C/EBPβ 和特異轉錄因子,如PPARγ和C/EBPα,共同募集KMT2D與增強子區域結合,催化H3K4—甲基化。(3)KMT2D促進CBP和/或p300的結合,活化增強子區域,使其標記乙酰化H3K27。轉錄因子和乙酰化的H3K27可募集bromodomain結構域結合蛋白4(BRD4),進而招募RNA聚合酶II,從而活化特異基因表達。
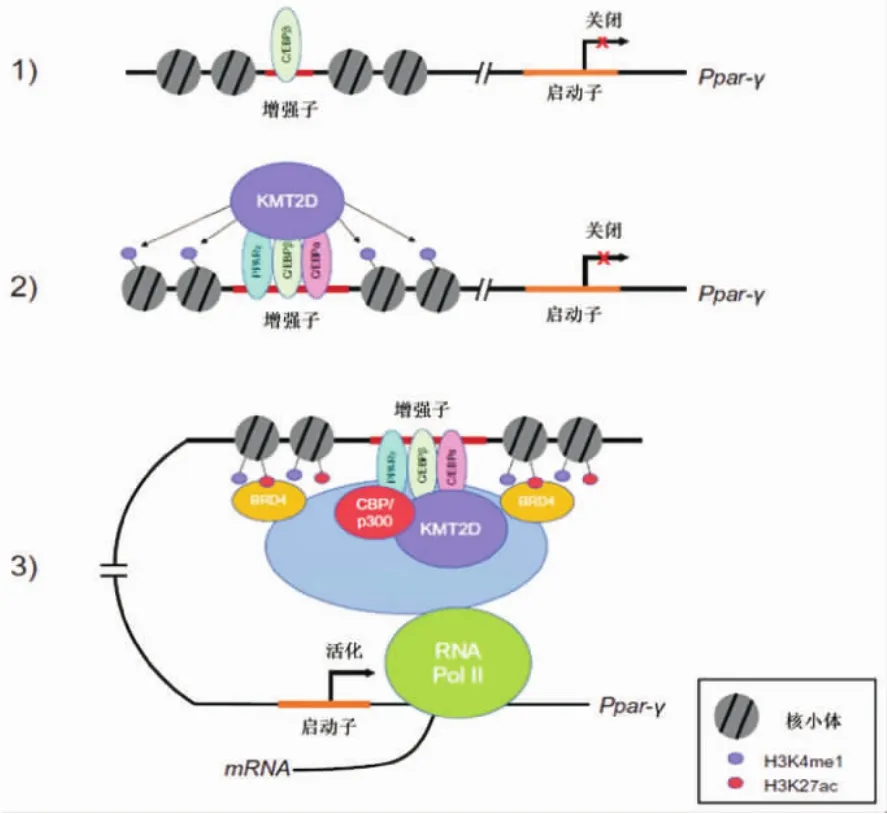
圖3 KMT2D調控Ppar-γ增強子,誘導特異基因表達的三步式模型
4 KMT2D在細胞發育、分化、新陳代謝和腫瘤抑制方面的作用
KMT2D在哺乳動物細胞中參與許多生命活動的進程,如發育、分化、新陳代謝和腫瘤抑制。其主要功能是通過激活增強子從而調控基因的表達。
4.1 發育和分化Kmt2d基因敲除的小鼠呈現出E9.5(胚胎第9.5 d)胚胎致死[13]。運用Myf5啟動子驅動的Cre轉基因小鼠,特異敲除肌肉和褐色脂肪前體細胞中的Kmt2d基因,導致褐色脂肪組織和肌肉質量的顯著下降,說明KMT2D在肌肉和脂肪組織發育中起重要作用[13]。特異敲除心臟前體細胞和心肌細胞中的Kmt2d基因導致嚴重的心臟缺陷和胚胎致死[17]。心臟發育進程中,KMT2D介導的H3K4me1/2在特異基因表達中有重要作用[17]。在B細胞發育過程中,KMT2D也扮演重要角色[18]。此外,KMT2D調控神經元和成骨細胞分化過程中特異基因的表達[4,19]。研究發現,KMT2D對于細胞命運轉變有重要作用,但對于胚胎干細胞和成體細胞的維持是非必須的[15]。
4.2 新陳代謝 在肝臟中,KMT2D與KMT2C也存在功能冗余,二者在多種新陳代謝過程中發揮顯著作用。Kmt2d+/-雜合小鼠,表現出糖耐受性、胰島素敏感性的增強,其血清膽汁酸水平升高[20]。Kim等[20]運用RNA-Seq分析發現,KMT2D和KMT2C對于肝臟生理節奏的表觀遺傳調控至關重要,作為生理節律轉錄因子視黃酸-相關孤兒核受體(ROR)-α和-γ的轉錄輔助因子發揮重要作用。該課題組實驗表明Kmt2d+/-小鼠表現出對高脂飲食誘導的肝脂肪變性的抵抗性[21]。
4.3 腫瘤抑制 大量研究表明,KMT2C和KMT2D具有腫瘤抑制作用。相關實驗表明,NCOA6和KMT2C或KMT2D作為抑癌基因p53轉錄輔助因子,是在DNA損傷藥物阿霉素作用細胞后,內源性p53基因表達所必須的[22]。小鼠體內研究表明,KMT2C和KMT2D在急性髓系白血病、濾泡性淋巴瘤和彌漫大B細胞淋巴瘤中起到抑制腫瘤的作用[23-24]。Chen 等[23]運用 RNAi和 CRISPR/Cas9 技術,研究發現KMT2C基因表達水平低于大約50%會誘發白血病。Zhang等[24]在Kmt2d敲除的小鼠中,過表達致癌基因Bcl2,導致生發中心來源的B細胞淋巴瘤的發生率上升。Ortega-Molina等[18]研究表明,敲除小鼠B細胞中的Kmt2d,會影響抑癌基因TNFAIP3,SOCS3和TNFRSF14的表達,并進一步誘發淋巴瘤。
另一方面,有研究發現,在乳腺癌和結腸癌中,KMT2D缺失會降低癌細胞增殖率[25-26]。此外,在雌激素受體陽性的乳腺癌細胞中,KMT2D活性升高會促進染色質解螺旋招募轉錄因子,如雌激素受體(ER)。在 MCF7細胞中,AKT結合并磷酸化KMT2D,使其甲基轉移酶活性和ERα活性降低[27]。研究發現,加入PI3K抑制劑使AKT失活,導致雌激素受體目的基因表達上調,會降低乳腺癌藥物的治療效果[27]。研究表明KMT2D在不同細胞類型中具有多種功能,這可能取決于基因活化過程中KMT2D招募的特殊的轉錄因子結合基因組的特異位點。
5 KMT2D突變導致發育疾病和腫瘤發生
已證明KMT2D基因突變是引發Kabuki綜合征最常見的原因。序列分析結果顯示,Kabuki綜合征患者中,KMT2D基因突變率高達56%~75%[28]。此外,先天性心臟病患者表現出大量的調節H3K4甲基化水平的基因的突變,其中包括KMT2D基因[29]。Ang等[17]發現,敲除小鼠Kmt2d基因將導致心臟發育缺陷。
KMT2D基因作為最常見的突變基因之一出現在多種癌癥中[30]。研究發現,在惡性腫瘤中,KMT2D基因的SET和PHD區域的突變,其中37%是移碼突變,60%是無義突變[7]。KMT2D基因突變導致多種組織發生癌變[7],如成神經管細胞瘤[31]、嗜鉻細胞瘤[32]、非霍奇金淋巴瘤[33]、彌漫性大B細胞淋巴瘤[34],食管鱗狀細胞癌[35]、胰腺癌[36]、前列腺癌[37]。經過對KMT2D分子在疾病發生發展中作用的不斷研究,已研發出其相應抑制劑,這可能為疾病的治療提供新的手段[7]。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
東方少年·布老虎畫刊(2023年8期)2023-08-01 15:45:12
科學大眾(2021年6期)2021-07-20 07:42:44
科學(2020年3期)2020-11-26 08:18:30
學苑創造·A版(2020年9期)2020-10-13 09:41:02
娃娃樂園·綜合智能(2019年3期)2019-04-03 09:17:36
中成藥(2018年2期)2018-05-09 07:19:34
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
湖北師范大學學報(自然科學版)(2015年2期)2016-01-10 08:41:55
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
