硒和谷胱甘肽復合作用對大麥苗生理和抗氧化特性的影響
2019-01-03 02:05:58趙鳳云宋新華
江蘇農業科學 2019年21期
趙鳳云 宋新華
摘要:用大麥苗分析單一硒(Se)、谷胱甘肽(GSH)和Se+GSH處理15 d對大麥苗富集Se與GSH及其抗氧化特性的影響。結果顯示,Se+GSH復合處理比單一Se或GSH處理增加了大麥苗對Se和GSH的積累量。抗氧化酶系統在Se+GSH復合處理與Se或GSH單一處理之間存在差異。Se+GSH復合處理的大麥苗過氧化氫酶(CAT)和谷胱甘肽S-轉移酶(GST)的活性高于Se或GSH單一處理,相反,過氧化物酶(POD)活性則低于Se或GSH處理。Se+GSH處理超氧化物歧化酶(SOD)的活性比GSH處理低,而抗壞血酸過氧化物酶(APX)活性則比GSH處理高。Se+GSH復合處理條件下谷胱甘肽還原酶(GR)的活性大于Se處理而小于GSH處理。Se+GSH處理的大麥苗丙二醛(MDA)含量無明顯變化,而總抗氧化能力(T-AOC)顯著提高。說明Se+GSH復合作用比單一GSH或Se更有利于促進GSH和Se的吸收與積累,同時增強抗氧化效果。
關鍵詞:硒;谷胱甘肽;復合作用;大麥苗;抗氧化特性
中圖分類號: S512.301文獻標志碼: A
文章編號:1002-1302(2019)21-0120-04
收稿日期:2018-08-23
基金項目:山東省自然科學基金(編號:ZR2015CL009);山東省淄博市科技發展計劃(編號:2017ZBXC196)。
作者簡介:趙鳳云(1963—),女,山東諸城人,博士,教授,主要從事植物生物學研究。E-mail:1795320202@qq.com。
大麥苗含有豐富的氨基酸、礦物質、活性酶、維生素等營養成分,具有良好的營養和保健作用,已經開發出大麥苗粉、大麥汁飲品、保健產品添加劑和大麥苗面類食品等保健食品,特別是富硒大麥產品的開發應用對提高人體免疫、預防缺硒引起的各種疾病至關重要[1-2]。硒是一些抗氧化酶和硒蛋白的重要組成成分,具有多種生物活性功能,它不僅是人體和動物必需的微量元素,而且對植物的生長、產量、品質、抗逆及抗氧化水平等都有重要的調節作用。對番茄、大蒜與胡蘿卜等植物的研究表明,適量的硒促進植物的生長,但過量的硒則抑制生長[3-5]。在水稻、蜈蚣草、番茄、鐵皮石斛和大麥等植物中研究發現,硒能提高還原型谷胱甘肽(GSH)與抗壞血酸(AsA)含量及抗氧化酶活性,減少過氧化氫(H2O2)的積累和丙二醛(MDA)的產生,緩解除草劑、重金屬、鹽和低溫對植物的傷害[6-10]。一定濃度范圍內外源硒使谷子產量、谷胱甘肽(GSH)、可溶性糖和粗蛋白含量增加,并提高過氧化物酶(POD)和超氧化物歧化酶(SOD)等抗氧化酶的活性[11-12]。GSH不僅是植物體內重要的抗氧化劑,也是人體抗衰老和增強免疫力不可缺少的活性分子,它是谷胱甘肽抗氧化系統的重要組成部分,在氧化防御反應、化合物代謝、核酸和蛋白質等大分子的生物合成、細胞信號傳導和蛋白質間相互作用等方面具有重要的調節作用[13-14]。GSH作為細胞內的重要還原劑,影響抗氧化系統中SOD、過氧化氫酶(CAT)、谷胱甘肽還原酶(GR)等多種酶的活性[14]。外源GSH影響植物抗氧化系統中保護酶的活性,改變內源GSH和AsA的含量,參與植物細胞對重金屬、極端溫度、干旱、高鹽等逆境脅迫的應答[15-16]。外源硒增加小麥、水稻、番茄等植物谷胱甘肽過氧化物酶活性和GSH含量,影響谷胱甘肽氧化還原循環[6,8,10,17]。硒和GSH系統在增強機體的抗氧化能力和預防過量自由基引起的疾病過程中發揮重要作用[1-2,18]。關于硒和GSH對生物生理生化的影響已開展了廣泛研究,但是對于這二者的復合作用則少見報道。本試驗旨在分析同時富集硒和GSH大麥苗生理生化特性的變化,為進一步開發富硒和GSH大麥苗功能食品奠定基礎。
1材料與方法
1.1試驗材料與處理
大麥種子用75%乙醇(30 s)和次氯酸鈉(15 min)消毒,無菌水沖洗干凈,置于培養盤放培養箱內26~28 ℃暗培養48 h,其間定時噴灑無菌水。然后將小苗移植到培養盆中(含滅菌的沙子,底部有濾網,外套無孔盆,便于換水),放在培養室[晝/夜相對濕度60%/80%,晝/夜溫度26 ℃/20 ℃,光照14 h,光照度200 μmol/(m2·s)]用霍格蘭(Hoagland)營養液培養3 d后開始處理。在營養液中分別或同時添加還原型谷胱甘肽(GSH)和亞硒酸鈉(Na2SeO3,Se),其中GSH每天遞增0.5 mmol/L,Se每天遞增0.1 μmol/L,直至最終濃度分別為GSH 3 mmol/L、Se 0.6 μmol/L,然后用終濃度培養15 d(每天換培養液),收割大麥苗用于測定各項指標。每種處理至少獨立重復3次,每次至少3個平行處理(約100株/盆)。
1.2試驗方法
1.2.1硒和GSH含量的測定
硒含量的測定依據GB 5009.93—2017《食品安全國家標準食品中硒的測定》中電感耦合等離子體質譜法(ICP-MS);GSH的提取和含量的測定參照Luwe等的方法[19]。
1.2.2酶液的提取及酶活性的測定
各種酶的提取參照Rao等的方法[20],APX的提取液額外加入5 mmol/L抗壞血酸。APX和POD活性的測定參照Rao等的方法[20];脫氫抗壞血酸酶(DHAR)活性的測定按照Chen等的方法[21];CAT活性的測定參照Rao等的方法[22];SOD活性的測定參照Zhao等的方法[23];GR和GST活性的測定參照Gronwald等的方法[24]。
1.2.3MDA含量的測定
MDA含量的測定參照林植芳等的方法[25]。
1.2.4大麥苗粉總抗氧化能力(T-AOC)的測定
總抗氧化能力的測定按T-AOC測定試劑盒(南京建成生物工程研究所)說明書進行。將各種處理的大麥苗用液氮研磨成粉,取真空冷凍干燥前/干燥后相同質量的大麥苗粉,按1 g ∶9 mL 的比例加入蒸餾水,冰水浴條件下制成勻漿液,4 247 r/min 離心10 min,用上清液測定。
以上試驗于2018年3—7月在山東理工大學生命科學學院植物分子實驗室和山東理工大學分析測試中心完成。
1.3數據處理與分析
試驗結果用3次獨立重復試驗的“平均值±標準差”來表示。用t-檢驗分析不同處理之間的差異,差異顯著水平用P<0.05或P<0.01表示。
2結果與分析
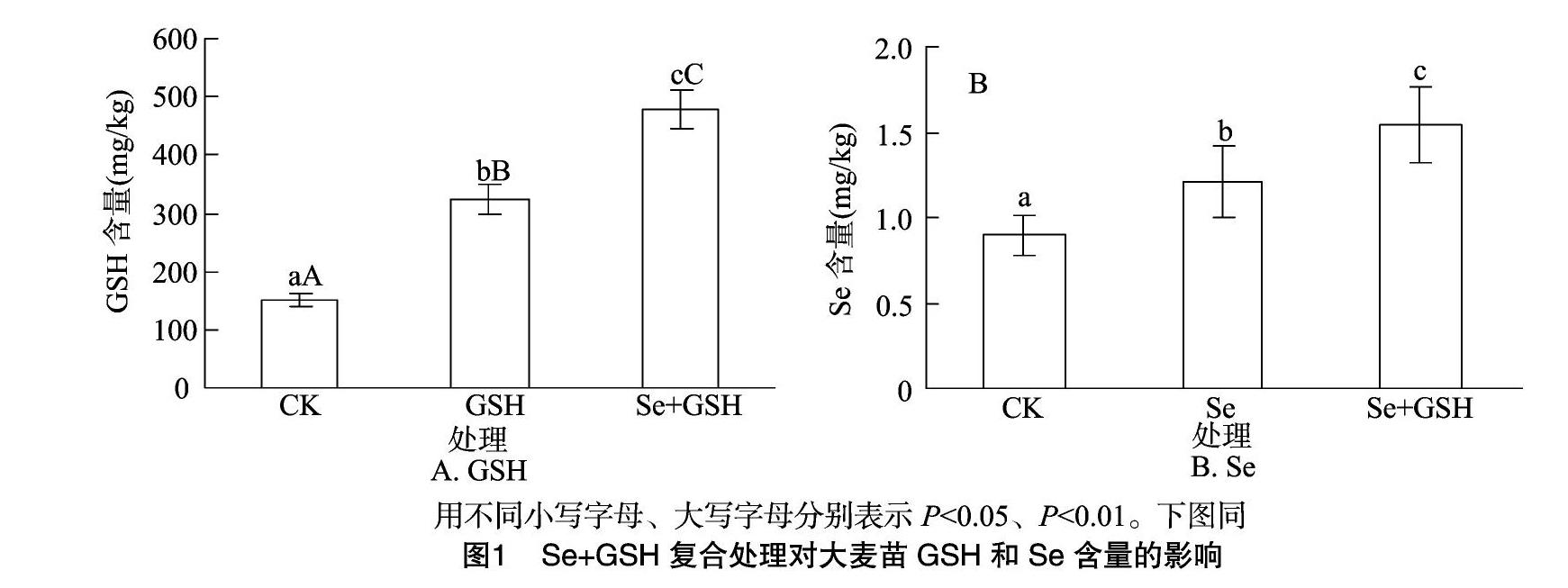
2.1Se+GSH復合處理對大麥苗GSH和Se含量的影響
如圖1-A所示,與對照比,單一GSH、Se+GSH處理的大麥苗GSH含量分別增加1.14、2.18倍,后者增加幅度大于前者,差異極顯著(P<0.01)。
如圖1-B所示,單一Se和Se+GSH處理的大麥苗Se含量分別比對照增加34.83%和72.02%,復合處理的Se含量比Se單一處理高27.57%(P<0.05)。
2.2Se+GSH復合處理對大麥苗抗氧化酶活性的影響
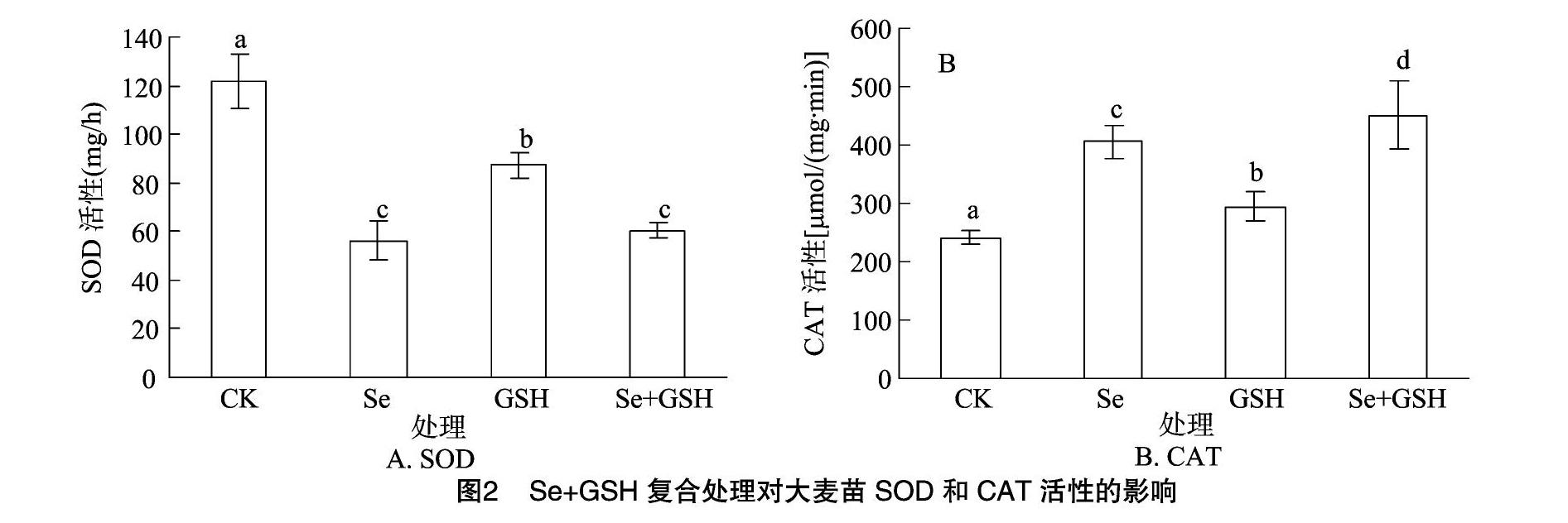
2.2.1Se+GSH復合處理對大麥苗SOD和CAT活性的影響
如圖2-A所示,與對照比,不同處理條件下大麥苗SOD活性都顯著降低(P<0.05)。其中,Se單一處理比對照低53.74%且與Se+GSH處理差異不顯著,但GSH單一處理的降低幅度小于Se和Se+GSH處理(P<0.05)。
如圖2-B所示,3種處理條件下的CAT活性都比對照有所提高(P<0.05),其中Se+GSH處理的大麥苗CAT活性增強程度最大,比對照高87.30%,比GSH單一處理高53.11%。
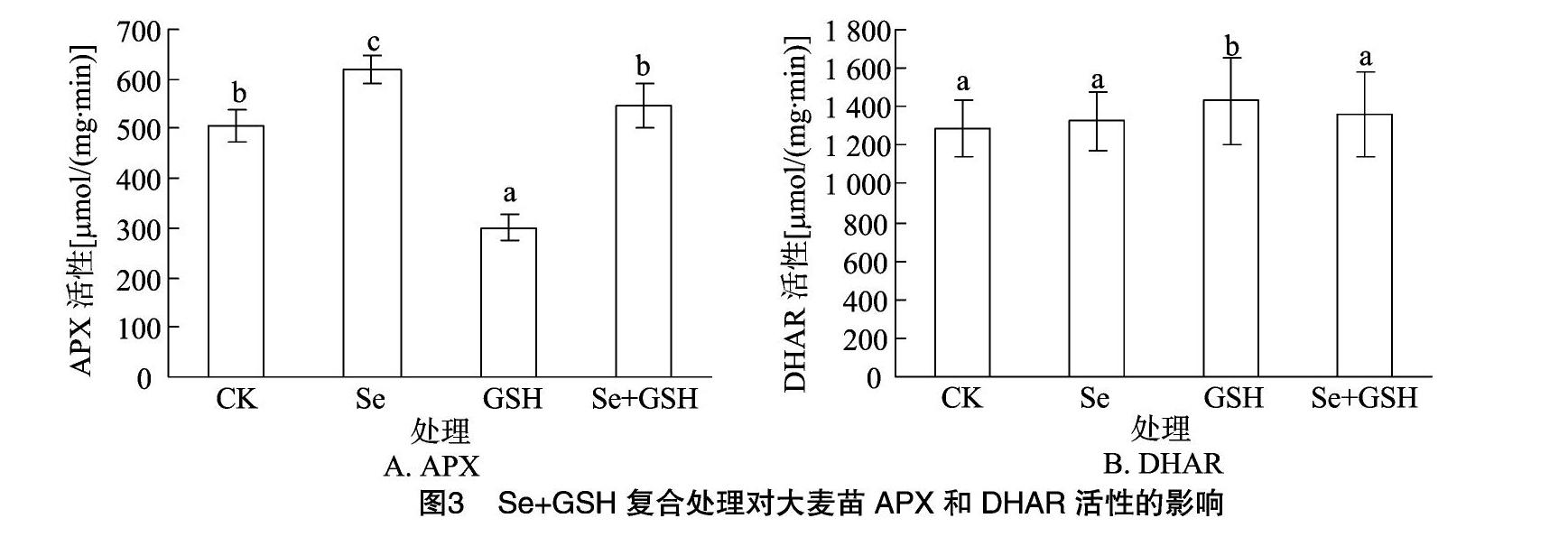
2.2.2Se+GSH復合處理對大麥苗APX和DHAR活性的影響
如圖3-A所示,3種處理條件下APX活性變化存在顯著差異,其中Se單一處理比對照高22.43%(P<0.05),GSH單一處理的APX活性比對照低40.52%(P<0.05),而Se+GSH 復合處理與對照差異不顯著。
如圖3-B所示,GSH單一處理的DHAR活性比對照增強11.26%(P<0.05),Se和Se+GSH處理的DHAR活性大小類似,且均與對照差異不顯著。
2.2.3Se+GSH復合處理對大麥苗GST和GR活性的影響
如圖4-A所示,3種處理的大麥苗GST活性都比對照強,其中Se+GSH處理的增加幅度最大,分別比對照、單一Se和單一GSH處理高107.38%(P<0.01)、84.48%(P<0.05)和
37.24%(P<0.05)。
如圖4-B所示,3種處理條件下的大麥苗GR活性都比對照低,但是降低程度有差異,其中,單一GSH處理的降低幅度最小,比對照低22.24%(P<0.05),其次是 Se+GSH復合處理(P<0.05),單一Se處理的GR活性最低,比對照低6946%(P<0.01)。
2.2.4Se+GSH復合處理對大麥苗POD活性的影響
如圖5所示,與對照比,單一Se處理的POD活性略有增強,但差異不顯著,單一GSH和Se+GSH處理的POD活性都減弱,其中Se+GSH復合處理的減弱幅度最大,比對照減弱47.53%(P<0.05)。
2.3Se+GSH復合處理對大麥苗MDA含量的影響
如圖6所示,各處理組MDA含量與對照組之間差異不顯著。
2.4Se+GSH復合處理對大麥苗粉總抗氧化能力T-AOC的影響
為了進一步分析Se+GSH復合處理大麥苗粉的抗氧化水平,分別測定真空冷凍干燥前后的大麥苗粉提取液總抗氧化能力T-AOC。如圖7所示,干燥前,對照和Se+GSH復合處理大麥苗粉的T-AOC分別為13.13、25.14 U/mL,后者的總抗氧化能力比前者高91.47%。真空冷凍干燥后,對照和Se+GSH復合處理的大麥苗粉的T-AOC都有所下降,與干燥前比,對照和復合處理的T-AOC分別降低21.48%和1679%,但復合處理的總抗氧化能力仍然比對照大1.03倍。
3討論與結論
硒在增強機體的抗氧化能力、預防缺硒和過量自由基引起的疾病過程中發揮重要作用[1-2,18]。通過植物吸收將有毒的無機硒轉化為無毒的有機硒是提高人體攝取硒的重要途徑。植物對硒的吸收積累能力與植物種類、發育時期、處理時間、處理方式、濃度及硒的價態等有關。對同一植物而言,在硒的價態和處理方式相同時,一定的時間和濃度范圍內,隨著時間的延長和濃度的增加植物對硒的吸收積累也增加,但是時間過長或濃度過高則會抑制植物生長并減少對硒的吸收[26-29]。本試驗中,萌發生長5 d的大麥苗用0.6 μmol/L亞硒酸鈉處理15 d時對硒的吸收積累效果較好。張承東等的研究表明,硒能提高GSH含量,影響抗氧化酶活性,減少H2O2的積累和MDA的產生,進而緩解逆境對植物的傷害[6-10]。GSH是谷胱甘肽抗氧化系統的重要組成部分,也是人體抗衰老和增強免疫力不可缺少的活性分子[13-14]。本試驗中Se+GSH復合處理條件下,大麥苗Se和GSH的含量比單一Se或GSH處理都多,說明Se+GSH復合作用促進了Se和GSH的積累。Se和GSH都能增強生物體的抗氧化能力,減少過量自由基的產生。抗氧化酶系統是植物體內清除自由基的重要途徑。本研究中,大麥苗幾個抗氧化酶的活性在Se+GSH 復合處理與單一Se或GSH處理之間存在差異。如Se+GSH處理SOD活性的減弱幅度大于單一GSH處理。就清除H2O2的抗氧化酶CAT、APX和POD而言,不同處理條件下其活性變化也不同,如Se+GSH處理的大麥苗CAT活性增強程度大于Se或GSH單一處理;Se+GSH復合處理的APX活性無顯著變化,而單一Se處理使APX活性增強,相反,單一GSH處理則使其活性減弱;Se+GSH復合處理的POD活性顯著弱于GSH或Se單一處理。GST和GR是谷胱甘肽抗氧化系統的重要酶,本試驗中Se+GSH處理的GST活性增強幅度顯著大于Se和GSH單一處理;無論是單一處理還是復合處理大麥苗GR的活性都減弱,但是單一GSH處理的減弱幅度小于單一Se和Se+GSH處理組的。本試驗條件下,Se+GSH處理的大麥苗MDA含量無明顯變化,而總抗氧化能力顯著提高。這些結果說明Se+GSH復合作用比單一GSH或Se更有利于促進GSH和Se的吸收與積累,同時增強抗氧化效果。本研究為進一步研發富集GSH和Se大麥苗功能營養粉奠定了重要基礎。
參考文獻:
[1]鄭慧敏,王軍,王夢竹. 大麥苗的營養價值及應用[J]. 大麥與谷類科學,2017,34(2):20-22,27.
[2]鮮瑤,張雷,宋戈,等. 大麥苗粉營養保健功能的研究進展[J]. 中國食物與營養,2016,22(11):73-76.
[3]施和平,張英聚,劉振聲. 番茄對硒的吸收分布和轉化[J]. 植物學報,1993,35(7):541-546.
[4]王永勤,曹家樹,李建華,等. 施硒對大蒜產量和含硒量的影響[J]. 園藝學報,2001,28(5):425-429.
[5]Biacs P A,Daood H G,Kodar I. Effect of Mo,Se,Zn and Cr treatments on the yield,element concentration,and carotenoid content of carrot[J]. Journal of Agricultural and Food Chemistry,1995,43(3):589-591.
[6]張承東,韓朔睽,魏鐘波. 硒對除草劑脅迫下水稻幼苗活性氧清除系統響應的作用[J]. 環境科學,2002,23(4):93-96.
[7]Srivastava M,Ma L Q,Rathinasabapathi B,et al. Effects of selenium on arsenic uptake in arsenic hyperaccumulator Pteris vittata L.[J]. Bioresource Technology,2009,100(3):1115-1121.
[8]王建偉,何曉玲,崔金霞,等. 外源硒對NaCl脅迫下加工番茄幼苗膜脂過氧化和AsA-GSH循環的影響[J]. 新疆農業科學,2014,51(10):1814-1820.
[9]張艷嫣,陳丹,譚艷玲,等. 外源硒對低溫脅迫下鐵皮石斛幼苗的緩解效應及其抗氧化生理特征變化[J]. 西北植物學報,2013,33(4):747-754.
[10]孫協平,羅友進,周廣文. 硒對甜櫻桃葉片褪黑素和谷胱甘肽氧化還原循環的影響[J]. 中國農業科學,2017,50(22):4373-4381.
[11]王永會,周大邁,張愛軍,等. 外源硒對谷子抗氧化酶活性及其品質的影響[J]. 中國土壤與肥料,2015(4):112-117.
[12]穆婷婷,杜慧玲,張福耀,等. 外源硒對谷子生理特性,硒含量及其產量和品質的影響[J]. 中國農業科學,2017,50(1):51-63.
[13]段喜華,唐中華,郭曉瑞. 植物谷胱甘肽的生物合成及其生物學功能[J]. 植物研究,2009,30(1):98-105.
[14]閆慧芳,毛培勝,夏方山. 植物抗氧化劑谷胱甘肽研究進展[J]. 草地學報,2013,21(3):428-434.
[15]Kocsy G,von Ballmoos P,Suter M,et al. Inhibition of glutathione synthesis reduces chilling tolerance in maize[J]. Planta,2000,211(4):528-536.
[16]劉傳平,鄭愛珍,田娜,等. 外源GSH對青菜和大白菜鎘毒害的緩解作用[J]. 南京農業大學學報,2004,27(4):26-30.
[17]郭靜成,尹順平. 硒對高等植物中谷胱甘肽過氧化物酶活性及谷胱甘肽含量的影響[J]. 西北植物學報,1998,18(4):533-537.
[18]Sies H. Strategies of antioxidant defense[J]. Pharmacological Research,1995,31(1):161.
[19]Luwe M W F,Takahama U,Heber U. Role of ascorbate in detoxifying ozone in the apoplast of spinach (Spinacia oleracea L.) leaves[J]. Plant Physiology,1993,101(3):969-976.
[20]Rao M V,Hale B A,Ormrod D P. Amelioration of ozone-induced oxidative damage in wheat plants grown under high carbon dioxide (role of antioxidant enzymes)[J]. Plant Physiology,1995,109(2):421-432.
[21]Chen Z,Young T E,Ling J,et al. Increasing vitamin C content of plants through enhanced ascorbate recycling[J]. Proceedings of the National Academy of Sciences of the United States of America,2003,100(6):3525-3530.
[22]Rao M V,Paliyath G,Ormrod D P,et al. Influence of salicylic acid on H2O2 production,oxidative stress,and H2O2-metabolizing enzymes. Salicylic acid-mediated oxidative damage requires H2O2[J]. Plant Physiology,1997,115(1):137-149.
[23]Zhao F Y,Wang X Y,Zhao Y X,et al. Transferring the Suaeda salsa glutathione S-transferase and catalase genes enhances low temperature stress resistance in transgenic rice seedlings[J]. Journal of Plant Physiology and Molecular Biology,2006,32(2):231-238.
[24]Gronwald J W,Plaisance K L. Isolation and characterization of glutathione S-transferase isozymes from sorghum[J]. Plant Physiology,1998,117(3):877-892.
[25]林植芳,李雙順,林桂珠,等. 水稻葉片的衰老與超氧化物歧化酶活性及膜脂過氧化作用的關系[J]. 植物學報,1984,26(6):605-615.
[26]付冬冬,段曼莉,梁東麗,等. 不同價態外源硒對小白菜生長及養分吸收的影[J]. 植物營養與肥料學報,2011,17(2):358-365.
[27]王家偉,陳雄波,黃起東,等. 不同水稻品種對硒的吸收轉化試驗[J]. 農業工程,2016,6(4):82-84.
[28]王曉潔,阮新,楊波,等. 大麥苗富硒研究[J]. 食品科學,2008,29(8):253-257.
[29]黃太慶,江澤普,邢穎,等. 水稻對外源硒的吸收利用研究[J]. 農業資源與環境學報,2017,34(5):449-455.
