不同水分條件對3種蕨類植物配子體形態發育的影響
2019-01-14 04:37:44高曉雯周施雨卞竹箐王清華戴錫玲
上海師范大學學報·自然科學版 2018年6期
高曉雯, 周施雨, 卞竹箐, 王清華, 戴錫玲
(上海師范大學 生命科學學院 植物種質資源開發協同創新中心,上海 200234)
0 引 言
我國蕨類植物資源豐富,約有2600種,占全世界蕨類總數的1/5.STOKEY[1-2]研究了多種蕨類植物配子體的發育過程,證實了其中的孢子萌發類型、配子體發育類型及成熟原葉體形態[3-5]在探討蕨類科、屬分類和系統演化方面具有一定的價值.但是,相對于種子植物,有些蕨類植物配子體性狀表現不穩定,易受環境條件影響而變化,例如,環境光的顏色、光照強度和溫度對蕨類植物孢子萌發、配子體生長、假根生長及兩性配子體的形成具有一定的影響[6-10].而水分條件對配子體形態發育的影響未見報道.
利用液體培養基培養烏毛蕨(Blechnumorientale)孢子[11],其孢子萌發時間較混合土培的明顯縮短,絲狀體長度更長,假根細長[12].用含水量較大的Knop′s瓊脂培養基和混合土對普通針毛蕨(Macrothelypteristorresiana)孢子進行培養[13],土培配子體在接種后35 d可觀察到精子器,而在含水量較大的Knop′s瓊脂培養基上的配子體產生精子器則需要60 d左右.采用MS(Murashige and Skoog)培養基、混合土和吸水濾紙分別培養傅氏鳳尾蕨(Pterisfaurieisp.)孢子[14],發現其配子體的形態與培養方式有關,MS培養基與土培的配子體假根粗短.以上研究結果表明,不同的水分條件可能是影響蕨類植物配子體形態發育的重要因素之一,對蕨類植物配子體形態中絲狀體形態、假根長短及性器官分化等存在一定影響.
本文依據PPG-1(2016)蕨類植物分類系統[15],選取薄囊蕨類(Polypodiidae)中莎草蕨目(Schizaeales)海金沙科(Lygodiaceae)海金沙[Lygodiumjaponicum(Thunb.) Sw.],水龍骨目(Polypodiales)中真水龍骨類(Eupolypods):真水龍骨類I(Eupolypods I)鳳尾蕨科(Pteridaceae)水蕨[Ceratopteristhalictroides (L.) Brongn],真水龍骨類II(Eupolypods II)烏毛蕨科(Blechnaceae)狗脊[Woodwardiajaponica(L.f.) Sm.]3種不同演化地位的蕨類植物,其中海金沙、水蕨配子體的形態發育已有報道[16-17].利用改良Knop′s瓊脂培養基,在其余環境條件完全相同的條件下,設置3組不同的水分條件,對3種蕨類植物配子體進行培養;利用光學顯微鏡對3種蕨類的孢子萌發、絲狀體、片狀體、原葉體、性器官和假根等形態特征進行詳細觀察,旨在進一步明確蕨類植物配子體形態發育過程中不受水分條件影響的穩定性狀,為蕨類植物配子體發育和分類演化研究提供基礎資料.
1 材料和方法
1.1 材 料
海金沙(L.japonicum)于2013年7月采自浙江寧波;水蕨(C.thalictroides)于2013年9月采自上海師范大學奉賢校區植物園;狗脊(W.japonica)于2013年8月采自浙江西天目山.將帶有成熟孢子的3種蕨類植物葉片置于潔凈紙袋中,放于干燥通風處,使孢子自然散落,約1周后將孢子去雜并收集于硫酸紙袋中.
1.2 方 法
在超凈工作臺上將5%(質量分數)次氯酸鈉溶液加入放有適量孢子的離心管中充分振蕩搖勻,浸泡5 min后離心(2000 r·s-1,3 min).去上清液,加入無菌水,振蕩離心.重復3次.在離心管中加入無菌水,配制孢子懸浮液.用膠頭滴管吸取1 mL孢子懸浮液,均勻涂布接種在平板培養基上(涂布密度小于100 mL-1),再按照無水、加入1管水(約2 mL)和2管水(約4 mL)分為3組,分別標記為組1、組2、組3,置于ZGX-300C型人工培養箱(浙江錢江儀器設備有限公司制造)中培養,全天光照,光照強度約為2000 lx,溫度為(25±2) ℃,濕度為60%~75%.
定期利用Nikon SMZ1500型光學顯微鏡觀察并記錄、照相.實驗重復3次,文中數據為3次觀察的平均值,用SPSS17.0軟件進行單因素方差分析.
2 結果與分析
2.1 不同水分條件下3種蕨類植物配子體的形態發育
2.1.1 海金沙配子體的形態發育
海金沙孢子接種后,組1、組2、組3的孢子萌發時間分別為16,14,11 d左右.海金沙孢子萌發類型為密穗蕨型(Anemia type)[18].
孢子萌發后形成絲狀體.比較了不同水分條件下海金沙配子體的發育形態,如圖1所示.由圖1可知:組1的絲狀體大多為單列細胞,長度為50~65 μm;組2和組3的絲狀體則具有單列細胞和雙列細胞兩種形態,且絲狀體長度為50~100 μm,明顯大于組1的長度.絲狀體頂端細胞經過多次縱裂,向兩側擴展,呈分散生長形成片狀體,片狀體進一步發育成心臟型原葉體,海金沙原葉體發育類型為鐵線蕨型(Adiantum type)[18].
當原葉體上產生性器官,表明原葉體已成熟.3個組都出現了雌性原葉體和雄性原葉體,雄性原葉體上精子器出現的時刻都早于雌性原葉體上頸卵器出現的時刻,兩種原葉體形態具有較大差異.海金沙接種約4周后,組3配子體首先出現精子器,約2 d后其余兩組也相繼出現精子器.精子器混生于假根叢中、兩翼下部及近中肋,近球形,直徑約為50~60 μm.3組產生的精子器的形態、大小無差異,且雄性原葉體的數量比3組雌性原葉體產生的數量多.海金沙接種9周左右后,組3最先出現頸卵器,頸卵器頂面觀為銅錢狀,且組3中雌性原葉體的數量多于組2和組1的.
海金沙配子體發育過程中,組1次生假根,始終粗短;組3次生假根細長;組2次生假根的長度介于組1和組3的長度之間.每組選取10個樣本進行測量,計算長度平均值,對組2和組1、組3和組1的次生假根長度進行顯著性分析比較,結果如表1所示,組3、組2的次生假根長度與組1的具有極顯著差異性(p<0.01).
2.1.2 水蕨配子體的形態發育
水蕨孢子接種后,組1、組2和組3的孢子萌發時間分別為7,6,5 d左右,水蕨孢子萌發類型為書帶蕨型(Vittaria type)[18].
接種9 d后,3個組絲狀體發育大小不同.比較了不同水分條件下水蕨配子體的發育形態,如圖2所示.由圖2可知,組2和組3的絲狀體長度明顯大于組1的,組1的絲狀體長度為(65.6±8.3) μm,組2的絲狀體長度為(114.7±38.6) μm,組3的絲狀體長度為(170.9±11.2) μm,絲狀體形狀均呈長條形.對組2和組1、組3和組1的絲狀體長度進行了顯著性分析比較,如上述結果所示,組3、組2絲狀體長度與組1具有極顯著差異性(p<0.01).
接種約12 d后,水蕨配子體進入片狀體階段,隨后發育成略不對稱的心臟型原葉體.水蕨原葉體發育類型為水蕨型(Ceratopteris type)[18].組1大多為雄性配子體,呈長條形;組2、組3大多為兩性配子體或雌性配子體.
水蕨接種約4周后,組3首先出現精子器,約2 d后其余兩組也相繼出現精子器.精子器的出現同樣早于頸卵器.精子器大多著生于原葉體邊緣;接種5周后,組3首先出現頸卵器,多發生于原葉體生長點下方的中脈范圍內.
組1的次生假根總體較短;組2的次生假根較長,組3的次生假根最長.對組2和組1、組3和組1的次生假根長度進行顯著性分析比較,由表1可知:組3、組2的次生假根長度與組1的具有極顯著差異性(p<0.01).

圖1 不同水分條件下海金沙配子體發育形態比較
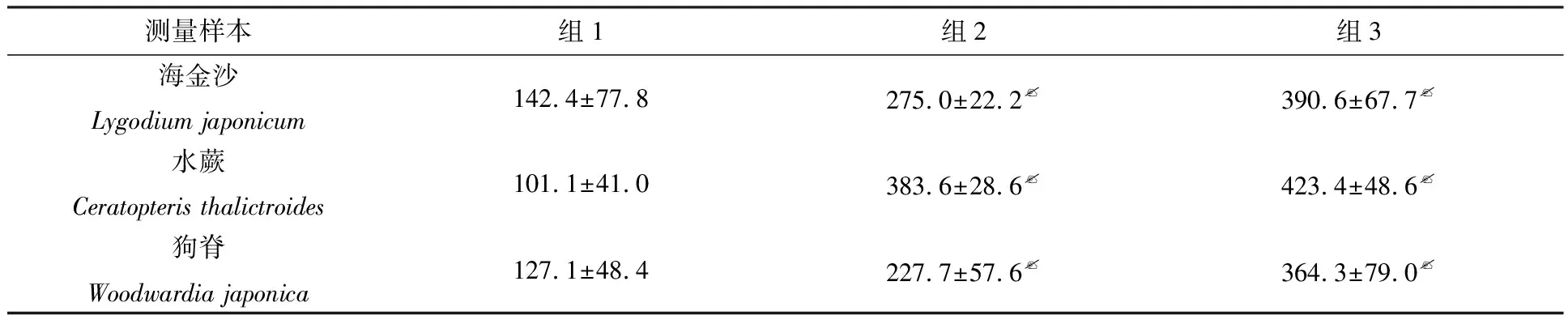
表1 不同水分條件下海金沙、水蕨和狗脊配子體次生假根長度比較 μm
注:p<0.01代表具有極顯著差異性

圖2 不同水分條件下水蕨配子體形態發育比較
2.1.3 狗脊配子體的形態發育
狗脊孢子接種后組1、組2、組3的孢子萌發時間分別為18,14,10 d左右,孢子壁從裂縫處裂開,長出初生假根以及含有葉綠素的原葉體原始細胞.狗脊孢子萌發類型為書帶蕨型(Vittaria type)[18].
原葉體原始細胞橫裂均形成單列絲狀體.比較了不同水分條件下狗脊配子體的發育形態,如圖3所示.由圖3可知,組1、組2絲狀體長度約為50~80 μm;組3絲狀體最長,為85~110 μm.絲狀體繼續發育,大多形成倒梯形片狀體.片狀體發育形成的原葉體基部微收,邊緣不平滑.在組3中明顯可觀察到:相隔幾個細胞便有一個細胞凸出,單細胞棒狀毛狀體長在突出細胞的頂端.組1大部分成熟原葉體展開時寬約800~1200 μm,組2的寬約1000~1500 μm,組3的寬約1000~1800 μm.狗脊原葉體發育類型為三叉蕨型(Aspidium type)[18].
狗脊配子體為兩性配子體.接種約6周后,組3首先出現精子器,約4 d后其余2組也相繼出現精子器.接種7周左右,組3首先出現頸卵器.
在狗脊整個配子體發育過程中,組1的次生假根粗短,組3的細長,組2的介于兩者之間.對組2和組1、組3和組1的次生假根長度進行了顯著性分析比較,由表1可知:組3、組2的次生假根長度與組1的都具有極顯著差異性(p<0.01).當配子體生長到成熟原葉體時期,組1的次生假根直徑約為20 μm,組2的約為17 μm,組3的與組2的較為接近.
各組成熟原葉體形狀沒有差別,但在水分充足時,原葉體邊緣毛狀體下方細胞分裂,毛狀體更突出,組1產生毛狀體數量最少,長度為(8.9±0.5) μm,一個原葉體約有5~15個毛狀體.組2的毛狀體長度為(18.3±0.9) μm,一個原葉體約有10~23個毛狀體.組3產生毛狀體數量最多,毛狀體長度為(20.3±1.1) μm,一個原葉體約有15~30個毛狀體,發育最發達.對3個組所測數據進行顯著性分析,結果顯示,組3、組2的毛狀體長度與組1的具有極顯著差異性(p<0.01),說明水分條件對毛狀體長度有影響.

圖3 不同水分條件下狗脊配子體形態發育比較
2.2 不同水分條件對3種蕨類植物配子體形態發育的影響
通過以上對比觀察,發現不同水分條件對3種蕨類植物配子體形態發育的影響表現在以下幾個方面:
1) 對孢子萌發速率的影響:海金沙、水蕨和狗脊的孢子萌發速率不盡相同,但均在水分最為充足時(組3)萌發最早,而在沒有水分時(組1)萌發最慢.
2) 對絲狀體長度的影響:在水分充足的條件下,3種蕨類植物形成的絲狀體長度最長,且海金沙在水分充足時易出現多列細胞的絲狀體.
3) 對配子體性別表現的影響:在缺水或水分不足條件下,3種蕨類容易形成雄性配子體,而在水分充足條件下,形成雌配子體的概率較大.
4) 對假根和毛狀體的影響:水分充足時,3種蕨類的配子體上次生假根更長,毛狀體更發達且數量更多;缺水時配子體上次生假根較短,毛狀體數量較少.
3 討 論
3種蕨類的孢子萌發類型及配子體發育類型不受水分條件的影響[16-17].在水分充足時,海金沙、狗脊和水蕨孢子萌發速率明顯較快,這與烏蕨(Stenolomachusanum)的情況相同[11-12];狗脊和水蕨的絲狀體長度更長,這與金毛裸蕨(Clematischrysocoma)、中華桫欏(Alsophilacostularis)和扇蕨(Neocheiropterispalmatopedata)的情況相同[7-8,10,16-17,19].另外,在海金沙組中還出現了進一步發育的雙列細胞絲狀體.在水分充足時,狗脊配子體產生毛狀體數量更多、發育更好,即毛狀體數量及發育程度會受水分條件影響[20],但毛狀體的形態、結構相對穩定[21].3種蕨類植物配子體的次生假根生長受水分條件影響較大[22],在水分不足的情況下,蕨類配子體次生假根基部膨大、末端分叉,而本實驗結果表明:蕨類配子體假根在水分充足時較長,在缺水時較短.觀察海金沙和水蕨的配子體發育發現:經歷不適條件或環境脅迫時,蕨類植物多發育為雄配子體[19,23];而在適宜條件下易發育形成雌性配子體或兩性配子體[24].這樣使蕨類植物在逆境條件下趨于產生大量精子,提高受精率,有利于物種的生存[8,24].
4 結 論
通過設置3個不同的水分梯度并控制其他環境影響條件,研究了海金沙、水蕨和狗脊3種不同演化地位的蕨類植物的配子體形態發育過程.結果表明:不同水分條件對不同演化地位的蕨類植物配子體發育形態都具有一定影響.在水分充足時,孢子萌發速率更快,絲狀體長度更長,性器官分化形成雌性配子體的概率更大,次生假根更長,毛狀體更發達且數量更多;在水分不足時,孢子萌發速率較慢,絲狀體較短,形成雄性配子體的概率更大,次生假根較短,毛狀體數量較少.若將上述特征作為蕨類植物分類鑒定的依據,需要嚴格控制其外部環境.孢子萌發類型、配子體發育類型、成熟原葉體形態及毛狀體形態結構不受水分條件的影響,是穩定的配子體特征性狀,可以作為蕨類植物分類演化研究的依據.
