亞溶解型C5b- 9刺激GMC誘導KLF4與PCAF結合及結合部位的鑒定
2019-01-25 03:45:14虞天一何風霞趙晨卉余明皓趙聃劉玉宮雅娟夏露邱文張婧王迎偉
東南大學學報(醫學版) 2018年6期
關鍵詞:實驗
虞天一,何風霞,趙晨卉,余明皓,趙聃,劉玉,宮雅娟,夏露,邱文,張婧,王迎偉
(1.東南大學附屬中大醫院 婦產科,江蘇 南京 210009; 2.南京醫科大學附屬第二醫院病理科,江蘇 南京 210003; 3.南京醫科大學 免疫學系,江蘇 南京 211166;4.南京醫科大學第一附屬醫院 腫瘤科,江蘇 南京 210029)
人類系膜增生性腎小球腎炎(mesangial proliferative glomerulonephritis,MsPGN)是以腎小球系膜細胞(glomerular mesangial cell,GMC)病變為主的腎炎[1],至今發病機制不明。大鼠Thy- 1腎炎(Thy- 1N)是研究MsPGN的模型,其病變有亞溶解型C5b- 9的依賴性[2- 3]。亞溶解型C5b- 9刺激GMC能上調一些轉錄因子和轉錄輔激活因子,調控基因表達,導致細胞凋亡[3- 4]或促炎因子合成[5],但其調控作用和機制尚未闡明。
Kruppel樣因子4(kruppel- like factor 4,KLF4)是一個Kruppel樣家族的轉錄因子,對細胞的發育和炎癥有調控效應[6]。P300/CBP相關因子(P300/CBP- assodated factor,PCAF)是具有乙酰轉移酶活性的轉錄輔激活因子[7],既能與轉錄因子作用,又可對蛋白進行乙酰化修飾[8],參與調控細胞周期和炎癥。
本實驗前期發現,在Thy- 1N大鼠腎組織中或體外,用亞溶解型C5b- 9刺激大鼠GMC后KLF4和PCAF表達上調,且兩者的表達時相較接近。鑒于亞溶解型C5b- 9是Thy- 1N大鼠GMC病變的啟動原,故本研究擬探討亞溶解型C5b- 9刺激GMC后KLF4與PCAF的結合情況及具體的結合部位,以期為今后解析MsPGN的致病機制提供基礎。
1 材料與方法
1.1 材料
大鼠GMC細胞系(HBZY- 1)由中國典型培養物保藏中心提供。多克隆兔抗Thy- 1抗體(Thy- 1 Ab)由本室制備。補體來自20名正常人血清(NS),補體熱滅活血清(heat- inactive serum,HIS)是指經56 ℃ 30 min滅活后的NS。人C6缺陷血清(human C6 deficient serum, C6DS)購于美國Complement Technology公司。重組人C6來自北京義翹神州生物技術有限公司。
免疫沉淀(immunoprecipitation,IP)細胞裂解液和兔IgG、羊IgG均購自上海碧云天生物技術研究所。小鼠β- actin單克隆抗體(BM0627)由武漢博士德生物工程有限公司提供。羊KLF4多克隆抗體(sc- 1905)購于美國Santa Cruz公司。兔PCAF單克隆抗體(ab- 176316)購于美國Abcam公司。
1.2 方法
1.2.1 GMC的培養和亞溶解型C5b- 9的確定 將GMC細胞系加入完全MEM液中培養。用棋盤滴定法即以不同濃度的Thy- 1 Ab致敏GMC,然后加不同濃度正常人血清孵育。之后,用乳酸脫氫酶(LDH)法測定GMC溶解百分率,GMC溶解壞死率小于5%視為亞溶解劑量[3- 5]。本實驗確定以5% Thy- 1 Ab與4% NS作為形成亞溶解型C5b- 9的最佳劑量。
1.2.2 亞溶解型C5b- 9刺激GMC時間點和分組處理 以亞溶解型C5b- 9分別刺激GMC 1、2、3、4、5、6、7和8 h;分成亞溶解型C5b- 9(5% Thy- 1 Ab+4% NS)、Thy- 1 Ab(5% Thy- 1 Ab)、Thy- 1 Ab(5% Thy- 1 Ab)+HIS(4% HIS)、Thy- 1 Ab(5% Thy- 1 Ab)+C6DS(4% C6DS)、Thy- 1 Ab(5% Thy- 1 Ab)+C6DS(4% C6DS)+C6(2 mg·L-1)和MEM培養液6組[4]。
1.2.3 質粒的構建、鑒定與轉染 構建pIRES2- KLF4和pIRES2- PCAF過表達質粒以及pIRES2- HIS- KLF4和pIRES2- HIS- PCAF結構域質粒,即(1)KLF4- S1(1~155 aa)、KLF4- S2(156~383 aa)、KLF4- S3(384~399 aa)和KLF4- S4(400~482 aa);(2)PCAF- S1(1~386 aa)、PCAF- S2(387~443 aa)、PCAF- S3(444~567 aa)和PCAF- S4(568~674 aa)。將上述質粒用NeonTM細胞電轉染系統轉入GMC中[3- 5]。
1.2.4 IP和IB實驗 不同處理的細胞加裂解液處理,具體的IP和IB實驗方法與步驟參見文獻[3- 5]。
1.3 統計學處理
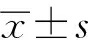
2 結 果
2.1 亞溶解型C5b- 9刺激GMC后KLF4與PCAF結合情況
用亞溶解型C5b- 9刺激GMC不同時間,結果顯示,在刺激2 h時KLF4與PCAF的結合明顯升高,3 h時達到峰值,見圖1。表明亞溶解型C5b- 9刺激GMC后使KLF4與PCAF的表達升高并相互結合。

A.電泳條帶;B.半定量分析,與MEM培養液組比較,aP<0.05
圖1亞溶解型C5b-9刺激GMC不同時間KLF4與PCAF結合情況
2.2 不同處理組的GMC中KLF4與PCAF相互結合水平的比較
將GMC按前述6組處理,用KLF4抗體IP后用PCAF抗體進行IB檢測。檢測結果發現,亞溶解型C5b- 9組和Thy- 1Ab+C6DS+C6組與KLF4結合的PCAF水平高于其他組,見圖2。提示亞溶解型C5b- 9刺激GMC 3 h確能提高PCAF與KLF4結合的水平。
2.3 KLF4、PCAF過表達質粒及KLF4、PCAF結構域質粒的構建與鑒定
IB檢測證實,轉染pIRES2- KLF4或pIRES2- PCAF的GMC中能表達KLF4或PCAF蛋白。將KLF4分成4個結構域,即KLF4- S1(1~155個氨基酸的轉錄激活結構域)、KLF4- S2(156~383個氨基酸的轉錄抑制結構域)、KLF4- S3(384~399個氨基酸的核定位信號

1為sublytic C5b- 9組,2為Thy- 1 Ab組,3為Thy- 1 Ab+HIS組,4為Thy- 1 Ab+C6DS組,5為Thy- 1 Ab+C6DS+C6組,6為MEM組,7為IgG對照組
A.電泳條帶;B.半定量分析,與MEM組比較,aP<0.01
圖2不同處理組GMC中KLF4與PCAF結合的比較
NLS)、KLF4- S4(400~482個氨基酸的鋅指DNA結合區域),并在5′端插入HIS標簽,構建了pIRES2- HIS- KLF4的4個結構域表達質粒。進而將PCAF蛋白分成4個結構域:分別為1~386個氨基酸的轉錄結合結構域、387~443個氨基酸的乙酰基轉移酶活性域、444~567個氨基酸的酵母轉錄輔激活子同源區、568~674個氨基酸的Bromo結構域,并命名為PCAF- S1、PCAF- S2、PCAF- S3和PCAF- S4。在5′端插入HIS標簽,構建了pIRES2- HIS- PCAF的4個結構域表達質粒。以上所有質粒測序結果正確,表明構建成功。
2.4 KLF4與PCAF結合部位的鑒定
將帶HIS的PCAF- S1、PCAF- S2、PCAF- S3和PCAF- S4分別與pIRES2- KLF4共轉染GMC 48 h,行IP和IB檢測發現,共轉染PCAF- S1和pIRES2- KLF4的KLF4表達量增高(圖3)。提示KLF4蛋白結合于PCAF的S1結構域(即轉錄結合結構域)。
將帶HIS的KLF4- S1、KLF4- S2、KLF4- S3和KLF4- S4分別與pIRES2- PCAF共轉染GMC后48 h,行IP和IB檢測證實,KLF4- S1與pIRES2- PCAF轉染組的KLF4表達量升高(圖4)。提示PCAF結合在KLF4的S1結構域(即轉錄激活結構域)。
3 討 論
KLF4是一種能參與轉錄調控的轉錄因子,其結構包括轉錄激活結構域、轉錄抑制結構域、核定位信號和DNA結合結構域。已有研究表明,KLF4能通過結合HMGB1調控炎癥因子的表達[6]。PCAF屬于一種轉錄輔激活因子,其結構含有轉錄結合結構域或PCAF同源結構域、乙酰基轉移酶活性域、酵母轉錄輔激活子同源區和布羅莫結構域。已有研究[9]發現,PCAF可與KLFs家族成員相互作用,并能乙酰化修飾KLF4,參與肺上皮的炎癥。

1為PCAF- S1+pIRES2- KLF4組,2為PCAF- S2+pIRES2- KLF4組,3為PCAF- S3+pIRES2- KLF4組,4為PCAF- S4+pIRES2- KLF4組
A.電泳條帶;B.半定量統計圖;與PCAF- S2+pIRES2- KLF4組、PCAF- S3+pIRES2- KLF4組、PCAF- S4+pIRES2- KLF4組比較,aP<0.01
圖3KLF4在PCAF上的結合部位鑒定

1為KLF4- S1+pIRES2- PCAF組,2為KLF4- S2+pIRES2- PCAF組,3為KLF4- S3+pIRES2- PCAF組,4為KLF4- S4+pIRES2- PCAF組
A.電泳條帶;B.半定量統計圖,與KLF4- S2+pIRES2- PCAF、KLF4- S3+pIRES2- PCAF組、KLF4- S4+pIRES2- PCAF組
圖4PCAF在KLF4上的結合部位鑒定
本實驗前期研究證實,在Thy- 1N大鼠的腎組織和GMC用亞溶解型C5b- 9處理后,KLF4和PCAF的表達明顯上調,且時相基本一致。因此設想,亞溶解型C5b- 9刺激GMC或許可誘導KLF4和PCAF相互結合,進而調控靶基因的轉錄。
為了確證這一推測,本實驗先用IP和IB鑒定了由亞溶解型C5b- 9刺激誘導的KLF4和PCAF的結合變化。結果顯示,在亞溶解型C5b- 9刺激GMC 2 h時KLF4和PCAF相互結合已明顯升高,3 h時達到峰值。進一步分組實驗表明,亞溶解型C5b- 9和Thy- 1 Ab+C6DS+C6組,KLF4和PCAF相互結合量也顯著上調。表明受亞溶解型C5b- 9刺激的GMC能誘導KLF4和PCAF結合增加。
本實驗又構建了帶HIS標簽的結構域表達質粒,分別與KLF4或PCAF的過表達質粒共轉染GMC。用標簽抗體行IP和IB檢測證實,KLF4結合于PCAF的轉錄結合結構域,而PCAF則結合在KLF4的轉錄激活結構域。提示在大鼠Thy- 1N時,在GMC膜上的亞溶解型C5b- 9可刺激GMC,誘導轉錄因子KLF4與轉錄輔激活因子PCAF相互結合形成復合物,而此復合物可能對其下游靶基因的轉錄發揮一定的調控作用。
猜你喜歡
作文·小學低年級(2025年2期)2025-02-13 00:00:00
小雪花·小學生快樂作文(2024年11期)2024-12-31 00:00:00
作文·小學低年級(2024年2期)2024-04-29 00:00:00
作文·小學低年級(2023年3期)2023-04-29 00:00:00
小獼猴智力畫刊(2022年9期)2022-11-04 02:31:42
小主人報(2022年4期)2022-08-09 08:52:06
中學生數理化·中考版(2022年11期)2022-02-16 07:01:20
小哥白尼(趣味科學)(2019年6期)2019-10-10 01:01:50
發明與創新(2016年38期)2016-08-22 03:02:52
太空探索(2016年5期)2016-07-12 15:17:55
