人ether-α-go-go鉀通道減少肝臟的內質網應激和凋亡
2019-01-28 05:55:22劉敬怡朱曉蓉謝榮榮袁明霞楊金奎
首都醫科大學學報 2019年1期
盧 晶 沈 涵 程 呈 劉敬怡 朱曉蓉 謝榮榮 袁明霞 楊金奎
(首都醫科大學附屬北京同仁醫院內分泌科 糖尿病防治研究北京市重點實驗室 北京市糖尿病研究所,北京 100730)
葡萄糖代謝是機體復雜的生理過程,包括胰島素的合成、成熟、分泌等環節。生理狀態下,胰島β細胞感受機體的血糖波動,分泌胰島素,促進肝臟、肌肉和脂肪的葡萄糖攝取,從而調節血糖至正常水平[1]。內質網是參與細胞內蛋白質折疊、成熟和轉運的重要細胞器,也參與細胞內鈣穩態及細胞死亡的調節過程。在缺氧、氧化應激或細胞內鈣穩態破壞等病理條件下,細胞為了自我保護,啟動未折疊蛋白反應(unfolded protein response,UPR)以應對內質網應激(endoplasmic reticulum, ER)[2]。UPR包括3條主要的信號轉導通路,分別由蛋白激酶RNA樣內質網激酶(protein kinase RNA-like endoplasmic reticulum kinase, PERK), 激活轉錄因子6(activating transcription factor 6, ATF6), 以及肌醇必需激酶1(inositol-requiring kinase1, IRE1)介導[3]。當內質網應激持續存在時,細胞可以誘發UPR的終末期反應——凋亡。已有研究[4-6]顯示,C/EBP-同源蛋白(C/EBP-homologous protein, CHOP)和IRE1下游通路參與細胞凋亡。腫瘤壞死因子受體相關因子2(tumor necrosis factor receptor-associated factor 2, TRAF2),凋亡信號調節激酶1(apoptosis signal-regulating kinase 1, ASK1)/ c-Jun氨基端激酶(c-Jun N-terminal kinase, JNK)以及半胱天冬酶半胱天冬酶12(cleaved-cysteine aspartyl protease-12,cleaved-caspase 12)的激活參與其中。內質網應激介導的肝細胞凋亡與肝臟損傷以及糖代謝異常關系密切。
生理狀態下,胰島β細胞的葡萄糖刺激的胰島素分泌(glucose-stimulated insulin secretion, GSIS)由一系列電活動調控,主要依賴于ATP敏感性鉀通道(ATP-sensitive K+channel, KATP)[7]。除KATP通道外,電壓門控ether-α-go-go相關基因(ether-α-go-gorelated gene, ERG)鉀通道家族也廣泛存在于胰島細胞。ERG家族屬于電壓依賴性鉀通道,有3個家族成員,分別是Kcnh2編碼的ERG1通道,Kcnh6基因編碼的ERG2通道以及Kcnh7編碼的ERG3[8]。在INS-1細胞中,ERG通道抑制劑E4031可以通過阻斷ERG通道,增加胰島素的分泌[9]。有研究[10]顯示,編碼的ERG1鉀通道參與心臟延遲整流電流。Hyltén-Cavallius等[11]的研究顯示,ERG1鉀通道突變的患者患有2型長QT間期綜合征且胰島素分泌升高。Rosati等[12]的研究顯示,ERG鉀通道在人胰島β細胞分泌胰島素的調節中具有重要作用。
然而,目前關于人ERG(human ERG, hERG)鉀通道和葡萄糖代謝關系的研究較少。本課題組發現一個hERG鉀通道雜合突變的糖尿病家系,家系中成年患者表現為低胰島素血癥和糖尿病,前期采用TALEN技術成功構建了hERG基因敲除小鼠動物模型,發現敲除動物存在糖耐量異常,但其具體機制尚未完全闡明[13]。本研究將利用課題組前期構建的hERG基因敲除動物模型進一步探討hERG基因敲除對存在肝臟內質網應激和凋亡的影響,以及這種影響是否導致了糖耐量異常。
1 材料與方法
1.1 實驗動物
SPF級健康的雄性C57BL/6J小鼠,8周齡,購于北京維通利華實驗動物技術有限公司。實驗動物許可證號:SCXK(京)2011-0012。采用TALEN技術構建hERG基因敲除小鼠動物模型(C57BL/6J背景)。實驗時,雄性hERG敲除小鼠(knockout, KO)與野生對照小鼠(wild type,WT)均為20周齡。飼養在環境溫度為20~24 ℃、環境濕度為50%~70%的SPF級動物房內,正常日光周期,自由進食及飲水,保持墊料干燥。
1.2 病理切片的制備
20周齡雄性小鼠,頸椎脫臼處死后,沿腹正中線打開腹腔,小心、迅速分離胰腺組織,將采集的胰腺置于4%(質量分數)多聚甲醛中固定24 h,以備制作石蠟切片。取出4%(質量分數)多聚甲醛固定24 h的胰腺和肝臟組織,梯度乙醇脫水,二甲苯中透明,將充分透明的胰腺組織塊放于融化的石蠟中,置于熔蠟箱內保溫。石蠟完全浸入組織塊后,進行包埋。將包埋好的胰腺組織蠟塊、肝臟組織蠟塊固定在切片機上,切成厚度為5 μm的薄片,薄片在預熱的水中燙平后貼于掛膠載玻片上,恒溫箱中烤干,以備染色用。
1.3 RNA的提取和熒光實時定量PCR
采用天根細胞組織總RNA提取試劑盒(貨號DP419)提取RNA,采用天根FastQuant cDNA第一鏈合成預混試劑(貨號KR108)合成cDNA。β-actin基因作為內參。采用Light Cycler 96儀器 (美國Roche公司)檢測RNA的表達情況。PCR反應體系為20 μL:2×酶混合物(Transgen Biotech公司, 貨號AS111) 10 μL、正向引物1 μL(10 μmol/L)、反向引物1 μL(10 μmol/L)、 cDNA 1.5 μL、DEPC H2O 6.5 μL。擴增條件為94 ℃ 5 min,94 ℃ 30 s,60 ℃ 30 s,72 ℃ 20 s,40個循環后再72 ℃ 7 min,自然冷卻至室溫。所用引物詳見表1。

表1 實驗用引物序列Tab.1 Primers used in the experiment
1.4 Western blotting
采用BCA蛋白分析試劑盒 (中國碧云天公司)測定蛋白濃度。取80 μg總蛋白進行10%(質量分數)的SDS-PAGE電泳,濕轉(200 mA,120 min)至硝酸纖維膜上。用TBST配制的5%(質量分數)的脫脂奶粉室溫封閉1.5 h,用封閉液將一抗稀釋后與膜在4 ℃搖床孵育過夜,次日用TBST洗膜3次,每次5 min,再與辣根過氧化物酶(horseradish Peroxidase, HRP)標記的二抗(1∶2 000)室溫孵育1 h。TBST洗滌,滴加增強化學發光(enhanced chemiluminescence, ECL)發光液后用Chemi-Doc Touch凝膠成像系統(美國Bio-Rad公司)顯影成像,并用Image J軟件對圖像進行灰度分析。所用抗體,詳見表2。

hERG: humanether-a-go-gorelated gene ;CHOP: C/EBP-homologous protein;caspase3: cysteine aspartyl protease 3;BIP(GRP78): binding immunoglobulin protein;ATF4: activating transcription factor 4;p-JNK: phosphorylated-c-Jun N-terminal kinase;JNK: c-Jun N-terminal kinase;Bax: Bcl2 associated X protein;Bcl2: B-cell lymphoma 2.
1.5 慢病毒感染
構建hERG的慢病毒LV載體。12周齡的雄性KO小鼠尾靜脈一次性注射慢病毒載體,慢病毒注射劑量為KO組:LV-hERG,WT組:LV-GFP,7×104 TU/g。兩周后處死小鼠,用Western blotting法檢測小鼠肝臟的內質網應激與凋亡的相關蛋白的表達水平。
1.6 統計學方法
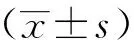
2 結果
2.1 hERG敲除小鼠肝臟形態學
小鼠20周齡時,分別檢測KO小鼠及其同窩WT小鼠的體質量及攝食量,二者差異均無統計學意義(圖1A、B)。小鼠頸椎脫臼處死后,分離小鼠肝臟,制備石蠟切片,行HE染色,顯微鏡下觀察。結果顯示,高倍鏡下,WT小鼠的肝細胞胞質豐富,胞核呈圓形或橢圓形,偶見雙核,肝血竇清晰可見,未見明顯的病理變化;KO小鼠的肝細胞輕度腫脹,胞質呈顆粒狀,肝血竇幾乎消失(圖1C)。
2.2 糖代謝關鍵酶在肝臟的表達
分離KO小鼠及同窩WT小鼠肝臟,Western blotting法檢測上述兩種糖異生關鍵酶在肝臟的表達,結果顯示與WT相比,葡萄糖-6-磷酸酶(glucose-6-phosphatase, G6pase)表達無明顯變化,磷酸烯醇式丙酮酸羧激酶(phosphoenolpyruvate carboxykinase, PEPCK)表達上調(P<0.01),提示hERG敲除可能引起肝臟糖代謝異常,詳見圖2、表3。

圖1 hERG敲除小鼠肝臟形態學Fig.1 Liver morphology of hERG knockout mice
A:weight of the mice;B: food intake of the mice;C: HE staining of the mice(100×);hERG: humanether-a-go-gorelated gene;WT: wide type;KO: knockout;n=6.

圖2 Western blotting檢測糖異生關鍵酶在小鼠肝臟的表達Fig.2 Expression level of the gluconeogenesis related key enzyme in the liver measured with Western blotting
* *P<0.01vsWT;WT: wide type;KO: knockout;G6pase: glucose-6-phosphatase;PEPCK: phosphoenolpyruvate carboxykinase.

表3 Western blotting法檢測糖異生關鍵酶在小鼠肝臟的表達水平Tab.3 Expression level of the gluconeogenesis related key enzyme in the liver measured with Western blotting
WT: wide type;KO: knockout;G6pase: glucose-6-phosphatase;PEPCK: phosphoenolpyruvate carboxykinase.
2.3 hERG敲除小鼠肝細胞凋亡增加
為研究肝臟變化的可能機制,對小鼠肝臟進行了凋亡相關指標的檢測。利用Western blotting的方法檢測肝組織中,凋亡相關蛋白的表達。結果顯示,與WT相比,KO小鼠肝臟的半胱天冬酶半胱天冬酶3(cleaved-cysteine aspartyl protease 3,cleaved-caspase 3),B細胞淋巴瘤因子2(B-cell lymphoma 2,Bcl2) 的表達上調和Bcl2相關X蛋白(Bcl2 associated X protein,Bax)表達下調,詳見圖3、表4。綜上,hERG敲除可以使小鼠肝細胞的凋亡增加。

圖3 Western blotting檢測小鼠肝臟凋亡相關蛋白表達Fig.3 Expression level of the apoptosis-related protein in the liver measured with Western blotting
*P<0.05vsWT group,n=3;WT: wide type;KO: knockout;p-JNK: phosphorylated-c-Jun N-terminal kinase;JNK: c-Jun N-terminal kinase;Bax: Bcl2 associated X protein;Bcl2: B-cell lymphoma 2;caspase3: cysteine aspartyl protease 3.
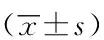
表4 Western blotting法檢測小鼠肝臟凋亡相關蛋白表達水平Tab.4 Expression level of the apoptosis-related protein in the liver measured with Western blotting
2.4 hERG敲除小鼠肝臟內質網應激水平增加
為進一步研究肝細胞凋亡是否與內質網應激有關,檢測了肝臟內質網應激相關蛋白的表達水平。real-time qPCR檢測結果顯示,KO小鼠的肝臟中結合免疫球蛋白的蛋白質(binding immunoglobulin protein, BIP)、CHOP、ATF4及ATF6的mRNA上調(圖4A)。Western blotting結果顯示,與WT小鼠相比,KO小鼠的肝臟中BIP(GRP78)、磷酸化真核細胞翻譯起始因子2(phosphorylated-eukaryotic translation initiation factor 2α, p-eIF2α)、ATF4以及CHOP的蛋白濃度上調(圖4B,表5)。綜上,hERG敲除小鼠的肝臟內質網應激水平上調。

圖4 小鼠肝臟內質網應激相關蛋白的表達Fig.4 Expression level of endoplasmic reticulum stress related protein in mouse liver using Western blotting
A: mRNA expression levels of stress related proteins in the liver using real-time PCR;B: protein levels of stress related proteins in the liver measured with Western blotting;*P<0.05,* *P<0.01vsWT group;hERG: humanether-a-go-gorelated gene ;WT: wide type;KO: knockout;BIP(GRP78): binding immunoglobulin protein;p-eIF2α:phosphorylated-eukaryotic translation initiation factor 2α;eIF2a: eukaryotic translation initiation factor 2α;ATF4: activating transcription factor 4;ATF6: activating transcription factor 6;CHOP: C/EBP-homologous protein.
WT: wide type;KO: knockout;hERG: humanether-a-go-gorelated gene;BIP(GRP78): binding immunoglobulin protein;eIF2a: eukaryotic translation initiation factor 2α;ATF4: activating transcription factor 4;CHOP: C/EBP-homologous protein.
2.5 hERG在體內過表達減輕肝臟內質網應激及凋亡
為進一步探究hERG在體內過表達能否改善 KO小鼠肝臟的內質網應激和凋亡,構建了hERG的慢病毒載體注射到實驗動物體內,兩周后處死小鼠,用Western blotting法檢測小鼠肝臟的內質網應激與凋亡的相關蛋白的表達水平。結果顯示與注射LV-GFP的對照組小鼠相比,LV-hERG組小鼠肝臟hERG表達明顯上調,顯示模型構建成功(圖5A)。LV-hERG組的ATF4及CHOP等內質網應激相關的蛋白表達水平下調,提示hERG過表達對小鼠肝臟內質網應激有保護作用。同時,Western blotting結果顯示,凋亡相關蛋白中,Bcl2的表達有上調趨勢(P=0.09),Bax、cleaved-caspase 3的表達下調,提示hERG過表達可改善小鼠肝細胞的凋亡(圖5B,表6)。綜上,hERG在體內過表達可以減輕肝臟的內質網應激和凋亡。

圖5 Western blotting法檢測小鼠肝臟內質網應激及凋亡相關蛋白的表達Fig.5 Expression levels of endoplasmic reticulum stress and apoptosis related proteins using Western blotting
A:hERG expression level of KO mice with LV-GFP or LV-hERG;B:expression levels of endoplasmic reticulum stress and apoptosis-related proteins;*P<0.05,* *P<0.01,** *P<0.001vsLV-GFP;hERG: humanether-a-go-gorelated gene;ATF4: activating transcription factor 4;CHOP: C/EBP-homologous protein;Bcl2: B-cell lymphoma 2.Bax: Bcl2 associated X protein;caspase3: cysteine aspartyl protease 3.
3 討論
肝細胞凋亡是導致肝臟損傷以及肝臟疾病的重要環節,內質網是真核細胞中參與細胞內跨膜蛋白與分泌蛋白合成、加工、修飾以及轉運的重要細胞器。內源性或外源性刺激引發的內質網蛋白折疊功能異常所導致的這種細胞應激狀態,稱為內質網應激。Ruiz-Vela等[14]的研究顯示,牛磺熊去氧膽酸可以通過抑制細胞內鈣離子的釋放而抑制內質網應激介導的肝細胞凋亡;可以降低胡蘿卜素誘導的細胞內鈣離子的上調,改善GRP78的過度表達,抑制Caspase12的激活,從而減輕肝細胞凋亡。
表6 Western blotting檢測小鼠肝臟內質網應激及凋亡相關蛋白的表達水平Tab.6 Expression level of endoplasmic reticulum stress and apoptosis-related proteins in the liver measured with Western blotting
肝臟在葡萄糖代謝,尤其是糖異生過程中發揮重要作用。糖異生是指乳酸、甘油以及生糖氨基酸等簡單的非糖前體,在一系列酶的催化作用下,轉變為葡萄糖或糖原的過程,它為機體血糖穩態提供了重要保證。G6pase是糖異生過程的關鍵酶,通過水解葡萄糖-6-磷酸鹽生成葡萄糖,在維持機體血糖穩態中發揮重要作用[15]。PEPCK是糖異生過程的另一種關鍵酶,在PEPCK作用下,將草酰乙酸轉化為磷酸烯醇式丙酮酸,經過一系列后續反應后最終生成葡萄糖[16]。本研究顯示,與同周齡WT小鼠相比,KO小鼠的糖異生關鍵酶PEPCK的表達上調,提示可能發生糖代謝異常,這一結果與課題組前期證實的KO小鼠存在糖耐量異常一致[13]。
KO小鼠肝臟內質網應激相關蛋白GRP78、p-eIF2α、ATF4和CHOP的轉錄和蛋白表達上調;凋亡相關蛋白cleaved-caspase3、Bax的表達上調,Bcl2表達下調。這一結果提示KO小鼠體內存在肝臟的內質網應激和凋亡。經尾靜脈向KO小鼠注射LV-hERG,Western bloting檢測結果顯示在KO小鼠體內,注射hERG病毒的小鼠有hERG基因表達,而注射GFP基因的小鼠體內沒有hERG表達,說明模型構建成功。結果顯示體內過表達hERG可以下調ATF4、CHOP、cleaved-caspase3等蛋白的表達。綜上,hERG可以通過改善肝臟的內質網應激與凋亡從而調節糖代謝。
本研究的局限性在于,沒有找到hERG與內質網應激反應的直接作用媒介以明確闡明hERG對內質網應激和細胞凋亡保護作用的機制。在后期的研究中,擬考慮通過免疫共沉淀等方法,尋找與hERG直接作用的分子。但可以明確的是,hERG對肝臟的內質網應激和凋亡具有保護作用,進而發揮改善糖代謝的作用。本研究可以為內質網鈣耗竭誘導的β細胞功能障礙的原因和hERG激動劑治療糖尿病的可能性提供新的見解。
猜你喜歡
興趣閱讀·興趣作文與閱讀(低年級)(2025年8期)2025-08-18 00:00:00
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2021年6期)2021-11-22 07:50:58
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
中學生數理化·七年級數學人教版(2020年12期)2021-01-18 06:57:46
學苑創造·A版(2020年9期)2020-10-13 09:41:02
小學生學習指導(低年級)(2017年10期)2017-10-10 01:00:05
海峽科技與產業(2016年3期)2016-05-17 04:32:12
云南中醫學院學報(2014年3期)2014-07-31 18:57:34
