棉花新品種冀豐1982 的高產機理
2019-02-07 08:06:54賈曉昀朱繼杰趙紅霞王士杰李妙王國印
河北農業科學 2019年5期
賈曉昀,朱繼杰,趙紅霞,王士杰,李妙,王國印
(河北省農林科學院糧油作物研究所,河北省作物遺傳育種實驗室,河北 石家莊 050035)
高產是棉花新品種培育的重要目標之一,挖掘高產品種的內在機理有助于提高新品種的選育效率,也可為田間高效管理提供技術支持[1]。產量形成是一個復雜、綜合的生理過程,協調好各方面之間的關系是實現作物高產的重要途徑[1,2]。較強的光合能力和光合產物輸出能力,較早的庫形成能力可為作物高產打下良好基礎[3]。光合能力的強弱可用葉面積指數、葉綠素含量、可溶性糖含量和可溶性蛋白含量等指標來表征[4,5],其中,可溶性糖還能夠促進植物較早地進入生殖生長,加快植物成熟[6]。衰老程度可用丙二醛(MDA) 含量、過氧化物酶(POD) 活性和超氧化物歧化酶(SOD) 活性等指標來表征[7]。通過以上指標可初步探究植物源庫關系以及產量形成的過程。
冀豐1982 為轉基因抗蟲常規棉品種,2014 年通過河北省審定。該品種長勢強、產量高。2011~2012年河北省冀中南春播常規棉花品種區域試驗2 a 平均子棉產量為3 882.0 kg/hm2,較對照冀棉958 增產6.69%,居參試品種第1 位;皮棉產量為1 549.5kg/hm2,較對照冀棉958 增產9.94%,居參試品種第2 位。以河北、山東、河南、天津、山西及國家黃河流域棉區區域試驗對照品種為參照,對冀豐1982 的生理生化指標進行分析,以期探究冀豐1982 的高產機理。
1 材料與方法
1.1 試驗材料
參試棉花品種有5 個,分別為冀豐1982(河北省農林科學院糧油作物研究所培育)、冀豐1271(河北省農林科學院糧油作物研究所培育)、冀棉958(河北省農林科學院棉花研究所培育)、中棉所41(中國農業科學院棉花研究所培育) 和魯棉研21(山東省棉花研究中心培育)。其中,冀棉958 和冀豐1271 為河北省區試對照品種,魯棉研21 為國家黃河流域棉區以及山東省、河南省和天津市區試對照品種,中棉所41 為國家黃河流域棉區以及山西省和河南省區試對照品種。
1.2 試驗方法
1.2.1 試驗設計 試驗于2015~2016 年連續2 a 在河北省農林科學院糧油作物研究所藁城堤上試驗站進行。以冀豐1271、冀棉958、中棉所41 和魯棉研21 為對照品種,冀豐1982 為試驗品種,設5 個品種處理。小區面積10.36 m2,3 行/區,行長7.4 m,行距0.7 m,株距0.32 m,隨機區組排列,3 次重復。其他管理同當地大田常規。
1.2.2 測定項目及方法 分別在苗期(6 月15 日)、蕾期(7 月4 日)、盛花期(7 月23 日)、盛鈴期(8月15 日) 和吐絮期(9 月4 日),每小區定行選取主莖功能葉(苗期和蕾期均取倒4 葉,盛花期取倒3 葉,盛鈴期取倒2 葉,吐絮期取倒1 葉) 5 片,測定葉片的葉綠素、可溶性糖、可溶性蛋白和MDA 含量,SOD 和POD 活性,以及葉面積指數。吐絮后,每小區隨機采摘50個正常吐絮的棉鈴,稱重,并計算單鈴重;軋花后,稱取皮棉重量,并計算衣分、子指。各小區均實收測產。
葉綠素含量測定采用95%乙醇浸泡法;可溶性糖含量測定采用硫酸-蒽酮比色法;可溶性蛋白含量測定采用考馬斯亮藍法;SOD 活力測定采用氮藍四唑(NBT) 光化還原法;POD 活力測定采用愈創木酚法;MDA 含量測定采用硫代巴比妥酸比色法。葉面積指數采用LAI-2200 型冠層儀測定。
1.2.3 數據分析 采用Excel 2010 和SPSS 17.0 軟件進行數據統計與分析。文中數據均為2 a 試驗的平均值。
2 結果與分析
2.1 冀豐1982 的產量及其構成因子
冀豐1982 皮棉產量最高,除與冀豐1271 差異不顯著外,與其他3 個對照品種的差異均達到了顯著水平;單株鈴數最多,除與冀棉958 差異不顯著外,與其他3 個對照品種的差異均達到了顯著水平;衣分含量中等,居參試品種第3 位;子指和單鈴重較高,均稍低于冀棉958,居參試品種第2 位。
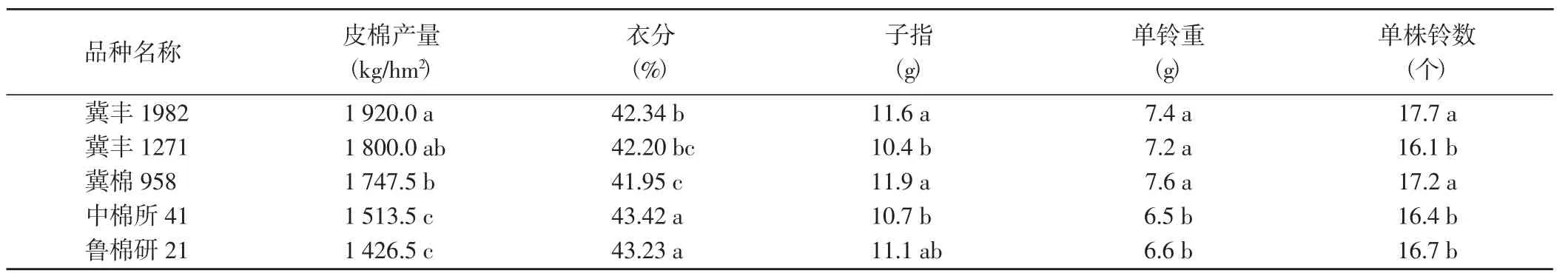
表1 參試棉花品種皮棉產量和產量構成因子的比較Table 1 Comparison of lint yield and yield components of tested cotton varieties
2.2 冀豐1982 的葉面積指數
苗期到吐絮期,5 個參試棉花品種的葉面積指數變化趨勢基本一致,均表現為先增加后降低,在盛鈴期達到最大值(圖1)。盛鈴期到吐絮期,中棉所41的葉面積指數下降最快,而冀豐1982 和冀棉958 的葉面積指數一直保持較高水平。
2.3 冀豐1982 的葉片葉綠素、可溶性糖和可溶性蛋白含量
苗期到吐絮期,5 個參試棉花品種的葉片葉綠素含量變化趨勢呈2 種類型:中棉所41 的葉片葉綠素含量表現為苗期到盛花期不斷增加,盛花期到吐絮期快速下降,生育后期葉片持綠能力弱于其他4 個品種;其他4 個品種的葉片葉綠素含量均表現為苗期到蕾期減少,蕾期到盛花期增加,盛花期到吐絮期緩慢下降,其中冀豐1982 的葉綠素含量一直高于其他3 個對照品種(圖2)。綜合而言,與對照相比,冀豐1982葉片的葉綠素含量一直保持較高水平,持綠能力最強。
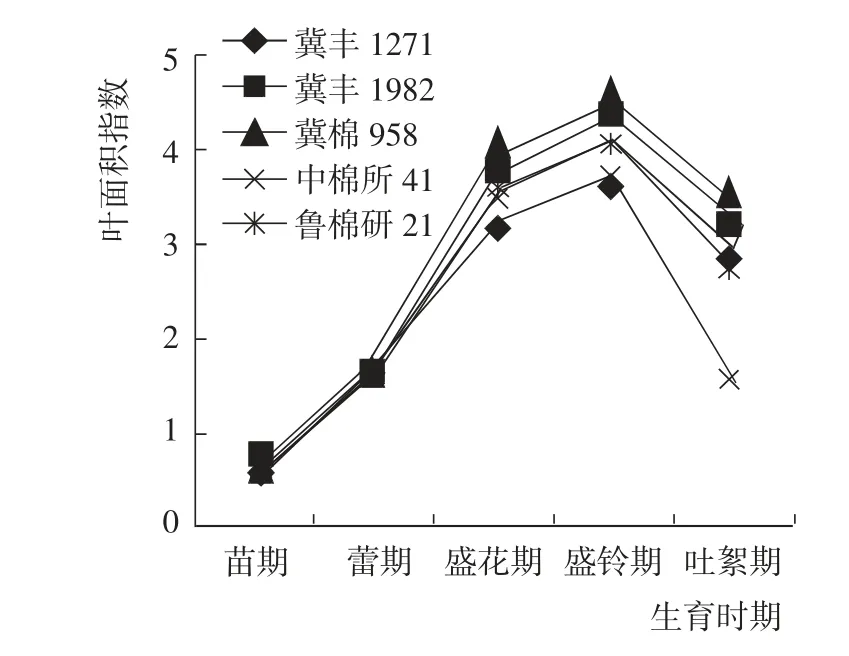
圖1 棉花生育期內葉面積指數的變化Fig.1 Change of leaf area index at different growth periods of tested cotton varieties
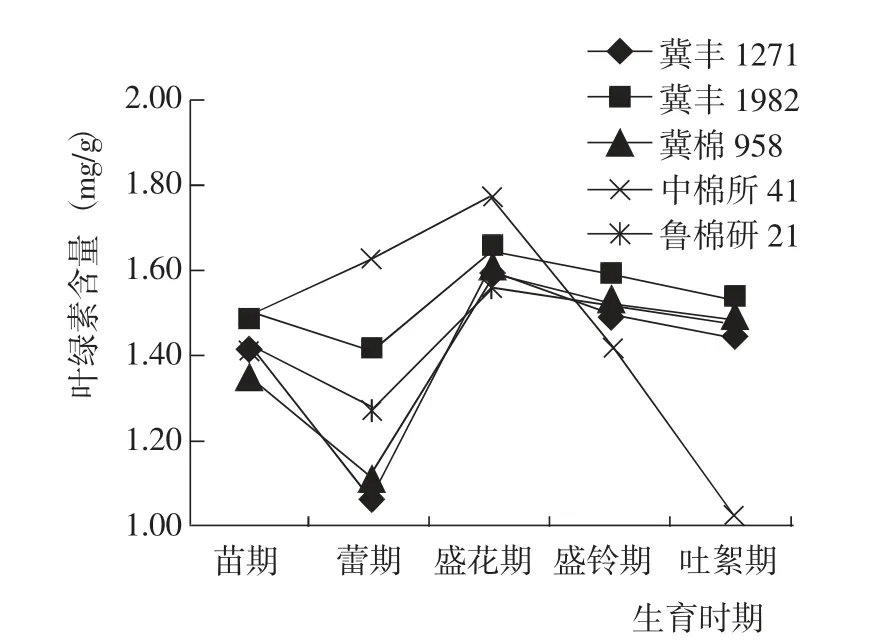
圖2 棉花生育期內葉片葉綠素含量的變化Fig.2 Change of chlorophyll content at different growth periods of tested cotton varieties
苗期到吐絮期,5 個參試棉花品種的葉片可溶性糖含量變化趨勢一致,均表現為增加—降低—增加—降低,在盛鈴期達到最大值,盛花期指標值最小(圖3)。苗期、盛鈴期和吐絮期,冀豐1982 的可溶性糖含量均最高;蕾期,冀豐1982 的可溶性糖含量僅次于中棉所41;盛花期,參試棉花品種的可溶性糖含量均較低,品種之間差異較小。綜合而言,冀豐1982 在苗期、蕾期和生育后期可溶性糖含量較高,有利于提早開花、結鈴和集中吐絮[13]。
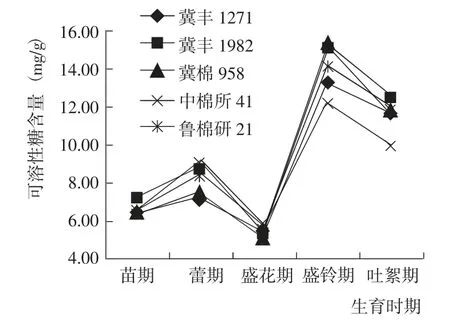
圖3 棉花生育期內葉片可溶性糖含量的變化Fig.3 Change of soluble sugar content at different growth periods of tested cotton varieties
苗期到吐絮期,5 個參試棉花品種的葉片可溶性蛋白含量變化趨勢基本一致,均表現為苗期到蕾期快速下降,蕾期到吐絮期緩慢波動下降(圖4)。苗期,冀豐1982 和冀棉958 的可溶性蛋白含量較高;蕾期和盛花期,冀豐1982 和冀豐1271 的可溶性蛋白含量較高;盛鈴期和吐絮期,5 個棉花品種的可溶性蛋白含量基本相當。綜合而言,在盛鈴期前,冀豐1982的可溶性蛋白含量較高;隨著葉片衰老,其可溶性蛋白含量平穩下降。與對照相比,冀豐1982 的葉片可溶性蛋白含量始終處于較高水平。
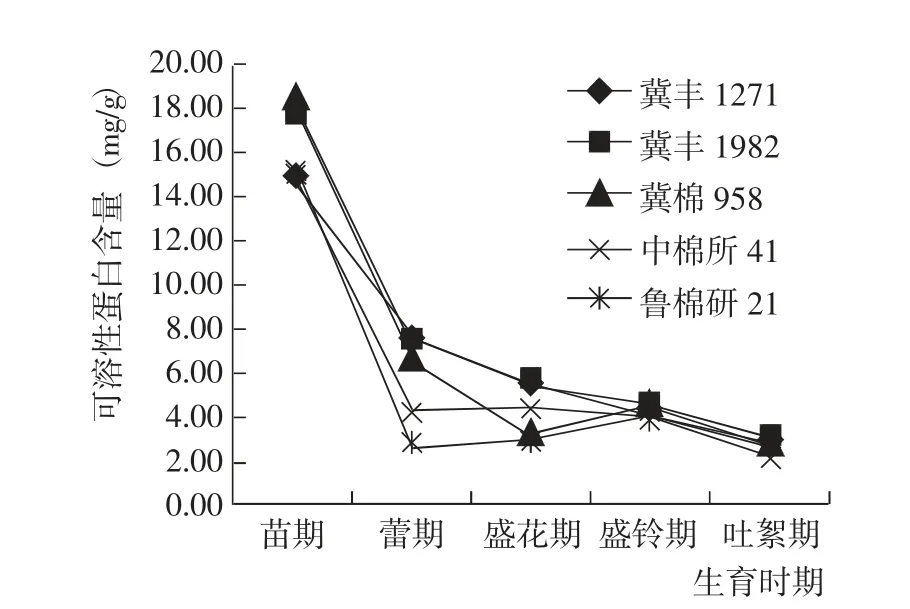
圖4 棉花生育期內葉片可溶性蛋白含量的變化Fig.4 Change of soluble protein content at different growth periods of of tested cotton varieties
2.4 冀豐1982 的葉片MDA 含量以及POD 和SOD活性
苗期到吐絮期,5 個參試棉花品種的葉片MDA 含量變化趨勢基本一致,均表現為增加—減少—增加,在盛花期達到最大值(圖5)。苗期和盛鈴期,5 個棉花品種的MDA 含量基本相當;蕾期和盛花期,冀棉958 和冀豐1982 的MDA 含量較低,其中盛花期指標值顯著低于其他3 個棉花品種;吐絮期,冀豐1982 和魯棉研21 的MDA 含量較低。綜合而言,與對照相比,冀豐1982 的葉片MDA 含量始終處于較低水平,衰老相對緩慢。
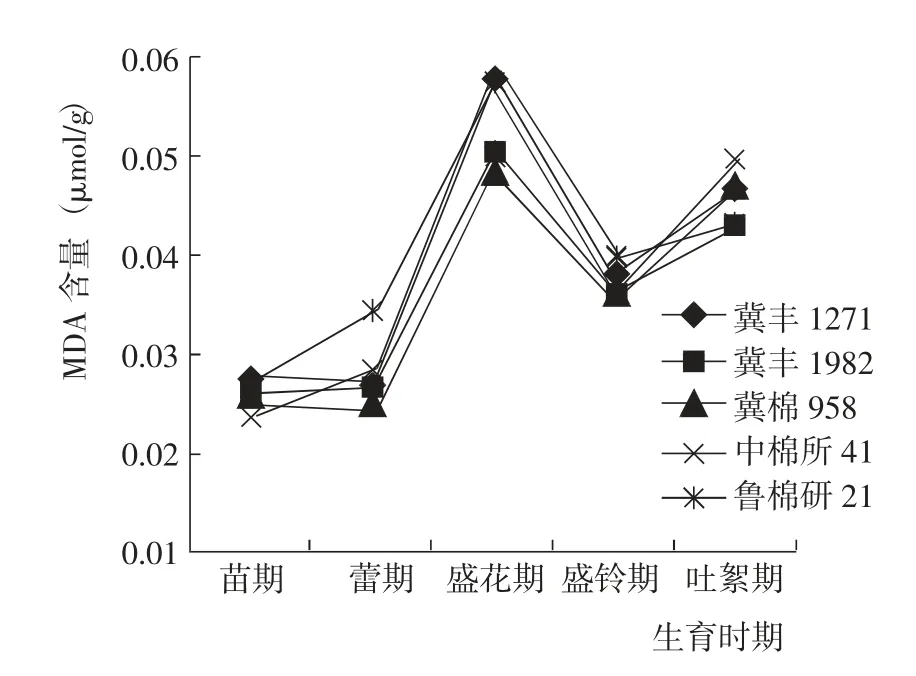
圖5 棉花生育期內葉片MDA 含量的變化Fig.5 Change of MDA content at different growth periods of tested cotton varieties
苗期到吐絮期,5 個參試棉花品種的葉片POD 活性變化趨勢一致,均表現為先增加后降低再顯著升高(圖6)。苗期到盛玲期,5 個棉花品種的POD 活性基本相當;吐絮期,各品種的POD 活性差異較大,其中,冀豐1982、冀豐1271 和冀棉958 指標值較高,魯棉研21 次之,中棉所41 最低。綜合而言,與對照相比,冀豐1982 的葉片POD 活性較高,具有較強的抗衰老能力。
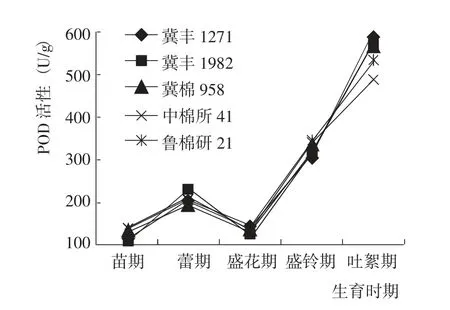
圖6 棉花生育期內葉片POD 活性的變化Fig.6 Change of POD activity at different growth periods of tested cotton varieties
苗期到吐絮期,5 個參試棉花品種的葉片SOD 活性均呈先降低后升高的變化趨勢,但變化節點及幅度有所不同(圖7)。其中,冀豐1982、冀豐1271 和冀棉958 的SOD 活性表現為苗期到蕾期快速下降,蕾期到盛玲期變化幅度較小,盛鈴期到吐絮期顯著升高,最終指標值達到或超過苗期水平;中棉所41 和魯棉研21 的SOD 活性表現為苗期到盛鈴期持續下降,盛鈴期到吐絮期開始升高,最終指標值未達到苗期水平。由此可見,冀豐1982 與冀豐1271 和冀棉958 在生長后期具有較強的抗衰老能力。
2.5 產量與各生理指標之間的相關分析
相關系數結果(表2) 顯示,蕾期、盛花期和盛玲期的葉面積指數均與子指呈極顯著正相關,吐絮期的葉面積指數與單鈴重呈顯著正相關;苗期的葉片可溶性蛋白質含量與子指呈顯著正相關;盛花期的葉片MDA 含量與子指和單鈴重呈顯著負相關;盛花期的葉片POD 活性與皮棉產量呈極顯著負相關;苗期的葉片SOD 活性與衣分呈極顯著正相關,與單鈴重呈顯著負相關。
綜上所述,光合能力相關指標正向顯著影響著棉花的子指和單鈴重;而衰老程度相關指標對皮棉產量、衣分、子指和單鈴重均有顯著影響,但是影響的方向正反不一,還有待進一步驗證。
3 結論與討論
棉花是我國乃至世界的重要經濟作物。2018 年我國棉花平均產量為1 818.3 kg/hm2,其中品種改良對單產提高的貢獻率達到30%以上[9]。棉花的4 個栽培種中,陸地棉對棉花總產量的貢獻達到95%以上[8],在棉花生產中占據主導地位。然而,隨著陸地棉遺傳多樣性的不斷減少,通過常規育種手段進一步選育產量高、綜合性狀優良的新品種愈加困難[1]。因此,通過研究高產品種的高產機理,可為棉花新品種選育開拓思路[1]。
有研究表明,棉花葉面積指數、葉綠素含量和可溶性糖含量增加能夠促進棉花干物質積累,減少棉鈴脫落,提高棉花產量[5,6]。生長后期葉片MDA 含量保持較低水平,POD 和SOD 保持較高活性并平穩下降,有利于棉花后期不早衰,實現高產、穩產[7]。
本研究中,以黃河流域棉區不同省份的4 個區試對照材料為參照,通過對比分析冀豐1982 的產量性狀以及部分生理指標,對冀豐1982 的高產機理進行了初步探究。冀豐1982 皮棉產量和單鈴數均居參試品種第1 位,單鈴重和子指均居參試品種第2 位,衣分居參試品種第3 位,各項產量指標均為中等偏上水平,相對協調。全生育期內葉面積指數、葉綠素含量、可溶性糖含量和可溶性蛋白含量較高,且后期能保持平穩下降;葉片MDA 含量相對較低,POD 和SOD 活性相對較高,能夠較好地保持后期不早衰,保證上部棉鈴正常成熟。綜合來看,全生育期內冀豐1982 的各項生理指標動態變化表現較好,能夠使植株持續保持較強的葉功能,從而獲得高產。
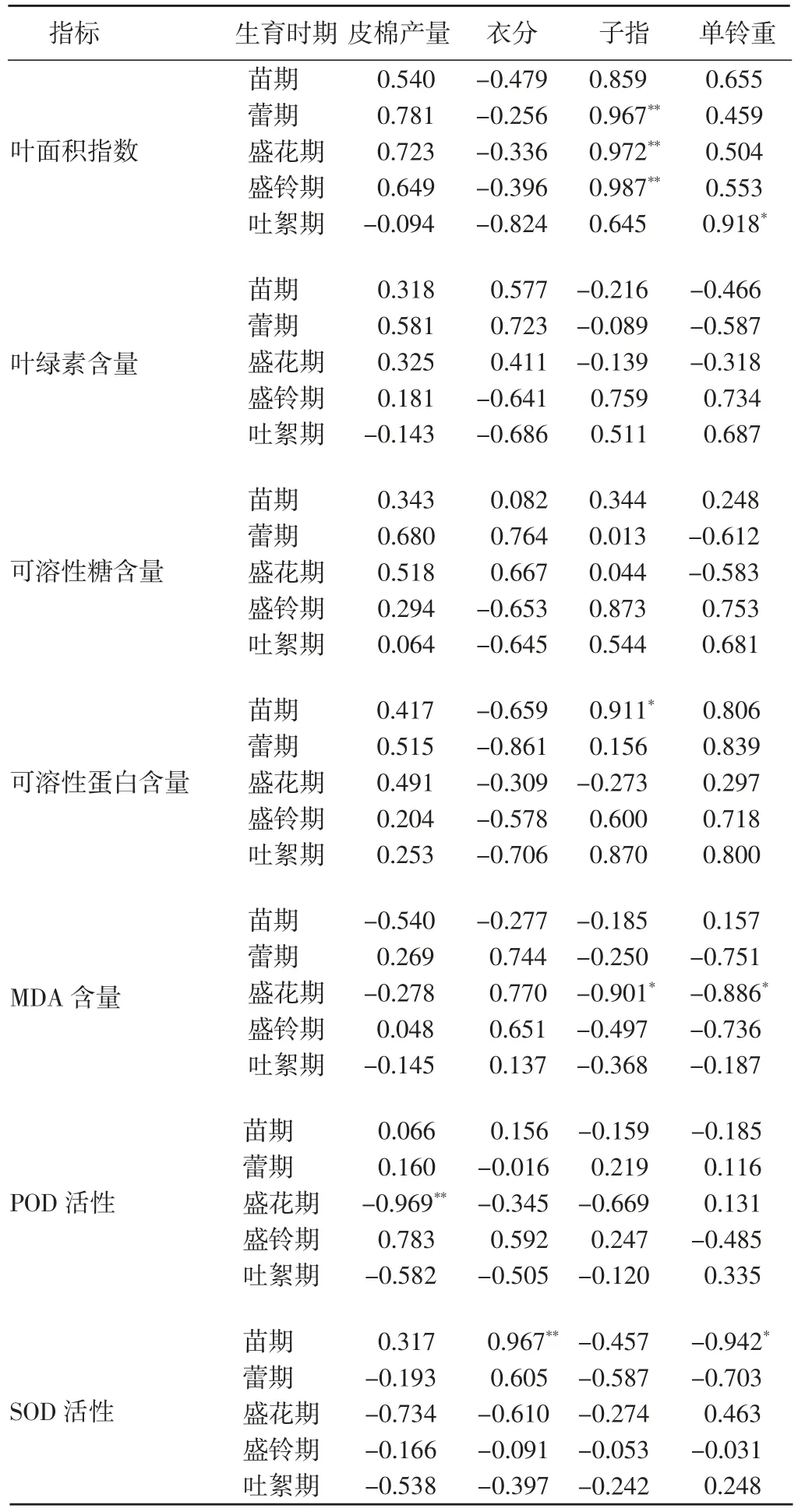
表2 產量及其構成因子與各生理指標的相關系數Table 2 Correlation coefficients among yield and its components with physiological indexes
隨著基因挖掘手段和技術的不斷進步,開展關鍵性狀的分子機理研究成為作物育種的重要方向。在棉花產量及其構成因子分子遺傳機制研究方面,已經發現了大量的相關遺傳位點,且部分基因功能得到了驗證[10~17],為棉花分子聚合育種提供了有用的標記基因資源,也為本研究提供了更多思路。下一步可結合冀豐1982 的生理研究,開展高產分子機制研究,以進一步挖掘其高產機理。
