草地貪夜蛾室內種群抗寒能力測定
2019-02-10 10:50:51張智鄭喬張云慧劉杰殷新田湯清波李靜袁源李祥瑞朱勛
植物保護 2019年6期
張智 鄭喬 張云慧 劉杰 殷新田 湯清波 李靜 袁源 李祥瑞 朱勛


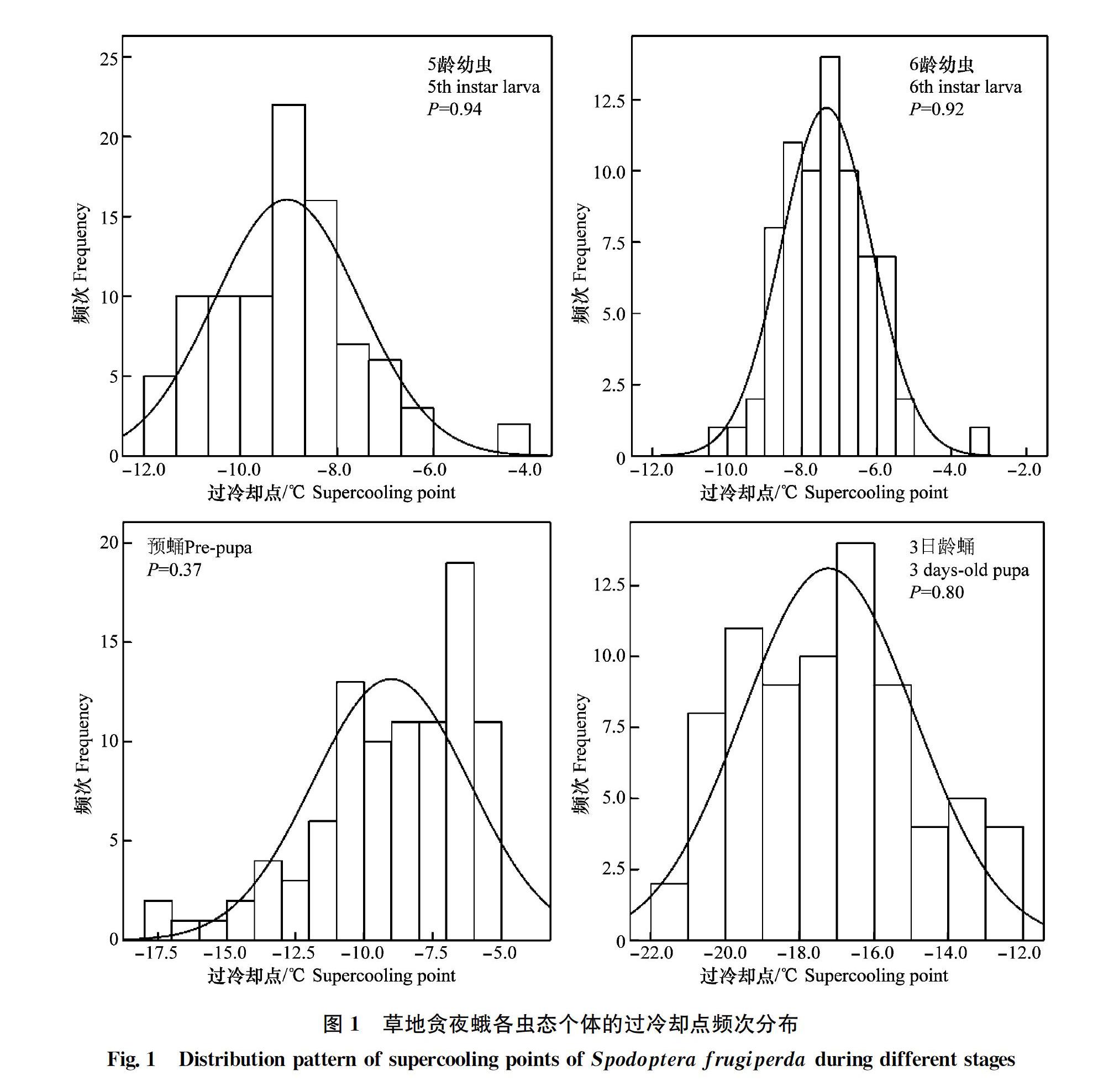
摘要 :為明確草地貪夜蛾的過冷卻點和結冰點,為判斷草地貪夜蛾抗寒能力提供依據,本研究利用過冷卻點測定儀對草地貪夜蛾不同蟲態的過冷卻點和結冰點進行了測定,并對其頻次分布進行了分析。結果表明:草地貪夜蛾的過冷卻點(df=834,F=430.26,P<0.01)和結冰點(df=834,F=891.71,P<0.01)在各蟲態之間存在極顯著差異,其中卵的過冷卻點和結冰點最低,分別為(-25.45±0.25)℃和(-24.70±0.25)℃,顯著低于其他蟲態。6齡幼蟲的過冷卻點和結冰點最高,分別為(-7.35±0.14)℃和(-2.07±0.11)℃;過冷卻點由低到高順序為:卵<1齡幼蟲<3日齡蛹<2齡幼蟲<1日齡成蟲<3齡幼蟲<4齡幼蟲<5齡幼蟲<預蛹<6齡幼蟲。結冰點的變化趨勢和過冷卻點相似,由低到高的順序為:卵<1 齡幼蟲<2 齡幼蟲<3日齡蛹<3 齡幼蟲<1日齡成蟲<4齡幼蟲<預蛹<5齡幼蟲<6齡幼蟲;幼蟲期,過冷卻點和結冰點隨齡期增加而逐漸升高。同一蟲態個體間的過冷卻點和結冰點頻次分布存在不同程度的變異。雌雄成蟲過冷卻點之間沒有顯著差異,雌成蟲結冰點顯著低于雄成蟲。本研究結果初步表明草地貪夜蛾具有較強的抗寒能力,該結論為草地貪夜蛾越冬區劃分和監測預警提供了科學依據。
關鍵詞 :草地貪夜蛾;?抗寒性;?過冷卻點;?結冰點
中圖分類號:
S 433.4
文獻標識碼:?A
DOI:?10.16688/j.zwbh.2019430
Cold hardiness of laboratory populations of Spodoptera frugiperda
ZHANG Zhi1,2,?ZHENG Qiao1,3,?ZHANG Yunhui1,?LIU Jie4,?YIN Xintian1,
TANG Qingbo5,?LI Jing3,?YUAN Yuan1,5,?LI Xiangrui1,?ZHU Xun1
(1. Institute of Plant Protection, Chinese Academy of Agricultural Sciences, Beijing?100193, China; 2. Beijing Plant
Protection Station, Beijing?100029, China; 3. Hebei Agricultural University, Baoding?071001, China; 4. National
Agro?Tech Extension and Service Centre, Beijing?100125, China; 5. Henan Agricultural University, Zhengzhou?450002, China)
Abstract
In order to clarify the cold resistance, and provide technical support for monitoring and early warning of fall armyworm (FAW)(Spodoptera frugiperda), the supercooling points and freezing points of all stages of FAW were measured using a thermocouple. It showed that the supercooling points and freezing points were significantly different among adult, egg, larva and pupa and the egg was of the lowest value, (-25.45±0.25)℃ and (-24.70±0.25)℃ respectively for supercooling point and freezing point. The 6th instar larvae showed the highest value, (-7.35±0.14)℃ for supercooling point and (-2.07±0.11)℃ for freezing point. The supercooling points from lowest to highest was egg<1st instar larva<3 days?old pupa<2nd instar larva<1 day?old adult<3rd instar larva<4th instar larva<5th instar larva Key words Spodoptera frugiperda;?cold hardiness;?supercooling points;?freezing points 草地貪夜蛾Spodoptera frugiperda(J. E. Smith)(也稱秋黏蟲 fall armyworm)隸屬于鱗翅目Lepidoptera夜蛾科Noctuidae,是聯合國糧農組織全球預警的重大遷飛性農業害蟲。草地貪夜蛾起源于美洲大陸,現已擴散至全球100多個國家,具有適生性強、遷飛距離遠、繁殖快、食性雜、為害隱蔽、防控難度大等諸多特點[12]。2019年1月,草地貪夜蛾首次入侵我國云南省普洱市江城縣,之后快速向我國糧食主產區擴散,其發展速度之快、波及范圍之廣非常罕見。截至8月17日,草地貪夜蛾在我國24個省份的1 366個縣(市、區)發生,全國發生面積達95.4萬hm2,并呈現繼續北擴態勢,新增發生區域呈條帶狀分布[3]。 昆蟲是一種變溫動物,環境溫度變化會引發新陳代謝等一系列生理生化反應。昆蟲對溫度變化的響應分為抗寒能力和高溫耐受能力,它們決定著昆蟲的分布與生活策略,是種群存在與發展的重要前提[45]。過冷卻點(supercooling point,SCP)是評價昆蟲抗寒能力高低的一種重要指標[67]。對褐飛虱、小地老虎、棉鈴蟲等害蟲的抗寒能力研究表明,明確其耐寒性對確定其分布范圍、越冬界限和發生趨勢等都起著至關重要的作用[810]。研究資料表明,在美國,草地貪夜蛾只在德克薩斯州和佛羅里達州的南部地區越冬,并且各蟲態在0℃以下均無法長時間存活[1112]。目前,草地貪夜蛾作為新入侵我國的重大遷飛性害蟲,其在國內的抗寒能力、越冬界限等仍是空白。本研究通過測定草地貪夜蛾各蟲態的過冷卻點和結冰點,初步明確其抗寒能力,以期為今后研究草地貪夜蛾在我國的潛在分布和相關預測預報提供基礎生物學依據。 1?材料與方法 1.1?供試蟲源 草地貪夜蛾蟲源為采自云南省普洱市江城縣(109°39′3″E, 22°40′53″N)冬玉米田的高齡幼蟲,化蛹、羽化后用5%的蜂蜜水飼養,收集卵塊在室內用新鮮玉米葉飼養繁殖1代。本次試驗供試蟲源為F3代種群,蟲態包括卵、各齡期幼蟲、預蛹、蛹(3日齡)和雌雄成蟲(1日齡)等。飼養條件為溫度(24±l)℃,濕度70%±5%、光周期L∥D=14 h∥10 h、光照強度18 000 lx。 1.2?試驗儀器 高低溫交變試驗箱GDW?150B(-40~150℃)由北京切克試驗設備有限公司生產,降溫速度1℃/min,本試驗最低溫度設置為-30℃。SUN?Ⅴ型智能過冷卻點測定儀及配套軟件由鵬程電子有限公司研發,40通路,數據精度為0.1℃,5 s記錄1次蟲體溫度變化,該儀器自動采集并以txt格式存儲數據,可用配套軟件實時繪制蟲體溫度變化曲線。 1.3?測定方法 測定前,從各蟲態草地貪夜蛾中隨機選取發育一致個體放入4℃冰箱10 h,取出后室溫恢復2 h以待測定。每蟲態測定60頭以上。測定準備工作分兩類,卵和1~2齡幼蟲為第一類,3齡以上幼蟲、預蛹、蛹和成蟲為第二類。卵和1~2齡幼蟲,單粒或單頭放置在寬4.5 cm、長6 cm的保鮮膜上,將過冷卻點測定儀的熱敏電阻感溫探頭與蟲體充分接觸后用保鮮膜把蟲體與探頭包好,并檢查兩者是否充分接觸;3齡以上幼蟲、蛹和成蟲置于1 mL離心管中,將熱敏電阻感溫探頭插入離心管使探頭與蟲體充分接觸。準備工作完成后將其置于高低溫試驗箱進行測試,當溫度降到一定值時,昆蟲體液開始結冰,此時的溫度即為過冷卻點,蟲體結冰后釋放出熱量,在溫度變化曲線上顯示為跳躍峰值,峰值即為結冰點[5]。 1.4?數據分析 數據分析軟件為IBM SPSS Statistics 19.0 for Windows。描述性統計分析主要計算了過冷卻點和結冰點的平均值和標準誤差,利用單樣本 Kolmogorov?Smirnov 法檢驗不同蟲態過冷卻點和結冰點的頻次分布特征。利用單因素方差分析(ANOVA)檢驗了不同蟲態對過冷卻點和結冰點的影響,并采用Duncan氏新復極差法進行了多重比較。分析性別對過冷卻點和結冰點的影響或比較兩種蟲態之間冷卻點或結冰點的差異時采用t檢驗。 2?結果與分析 2.1?不同蟲態的過冷卻點 草地貪夜蛾的過冷卻點在不同蟲態之間存在極顯著差異(df=834,F=430.26,P<0.01)(表1)。過冷卻點由低到高的順序依次為:卵<1齡幼蟲<3日齡蛹<2齡幼蟲<1日齡成蟲<3齡幼蟲<4齡幼蟲<5齡幼蟲<預蛹<6齡幼蟲。卵的過冷卻點最低,為(-25.45±0.25)℃,其次為1齡幼蟲,為(-19.02±0.41)℃。3日齡蛹和2齡幼蟲的過冷卻點也較低,分別為 (-17.23±0.27)℃和(-16.70±0.41)℃,但之間差異不顯著(df=139,t=-1.11,P=0.27)。4齡幼蟲、5齡幼蟲和預蛹的過冷卻點比較接近,6齡幼蟲過冷卻點最高,達(-7.35±0.14)℃(表1)。在幼蟲階段,隨著齡期的增加過冷卻點也逐漸升高(表1)。達到預蛹階段,過冷卻點開始降低。進入蛹期,過冷卻點進一步降低。成蟲羽化后,過冷卻點開始略微升高。預蛹、3日齡蛹和1日齡成蟲的過冷卻點之間存在極顯著差異(df=297,F=170.65,P<0.01)。 2.2?不同蟲態的結冰點 草地貪夜蛾的結冰點在各蟲態之間存在極顯著差異(df=834,F=891.71,P<0.01)(表1)。各蟲態結冰點由低到高的順序依次為: 卵<1 齡幼蟲<2 齡幼蟲<3日齡蛹<3 齡幼蟲<1日齡成蟲<4齡幼蟲<預蛹<5齡幼蟲<6齡幼蟲。卵的結冰點最低,為(-24.70±0.25)℃,6齡幼蟲的結冰點最高,為(-2.07±0.11)℃,個體的極差值可達27.26℃。幼蟲階段,結冰點隨著齡期的增加逐漸升高,最高可達-0.37℃。進入化蛹階段后,結冰點的變化趨勢與過冷卻點類似,也為先降低再升高。預蛹、3日齡蛹和1日齡成蟲的結冰點之間存在極顯著差異(df=297,F=169.56,P<0.01)。 2.3?不同蟲態過冷卻點和結冰點的頻次分布 草地貪夜蛾各蟲態個體的過冷卻點存在不同程度變異(圖1),其分布范圍具有一定寬度。過冷卻點最低值為卵期-28.44℃,最高值出現在6齡幼蟲,為-3.25℃。1齡幼蟲的過冷卻點分布范圍最寬為-10.03~-24.7℃,4齡幼蟲的過冷卻點分布范圍最小為-6.45~-12.73℃。過冷卻點分布寬度范圍由大到小依次為:1齡幼蟲(14.67℃)>2齡幼蟲(14.32℃)>1日齡成蟲(14.08℃)>預蛹(12.32℃)>卵(10.42℃)>3日齡蛹(9.08℃)>3齡幼蟲(8.14℃)>5齡幼蟲(7.73℃)>6齡幼蟲(6.89℃)>4齡幼蟲(6.28℃)。除卵的過冷卻點頻次不符合正態分布以外(P<0.01),其余各蟲態過冷卻點頻次均服從正態分布(P >0.05)。 草地貪夜蛾各蟲態個體的結冰點和過冷卻點相似,也存在不同程度變異(圖2),冰點最低值為卵期-27.63℃,最高值為預蛹期-0.37℃。3日齡蛹的結冰點分布范圍最寬為-3.85~-20.37℃,6齡幼蟲結冰點分布范圍最小為-0.92~-6.66℃。具體寬度范圍由大到小為:3日齡蛹(16.52℃)>1齡幼蟲(15.23℃)>2齡幼蟲(14.55℃)>3齡幼蟲(11.3℃)>4齡幼蟲(10.66℃)>1日齡成蟲(10.40℃)>卵(10.35℃)>5齡幼蟲(9.21℃)>預蛹(8.60℃)>6齡幼蟲(5.74℃)。卵、5齡幼蟲、6齡幼蟲、預蛹的結冰點頻次不符合正態分布(P<0.05),其余蟲態結冰點頻次均服從正態分布(P>0.05)。 2.4?雌雄成蟲的過冷卻點和結冰點 雌成蟲過冷卻點為(-15.11±0.45)℃,雄成蟲的過冷卻點(-14.93±0.44)℃,兩者之間差異不顯著(df=125.29,t=-0.29,P=0.773)。雌成蟲結冰點為(-6.19±0.23)℃,雄成蟲的結冰點為(-5.46±0.19)℃,兩者之間差異顯著(df=126,t=-0.2.37,P=0.02<0.05)。 3?討論 過冷卻點代表昆蟲能夠存活的生理低溫下限,是反映昆蟲越冬抗寒性的一個主要指標。過冷卻點越低,昆蟲抗寒性越強[13]。本文對草地貪夜蛾各蟲態的抗寒性測試結果顯示卵的過冷卻點和結冰點最低,其次為蛹和成蟲,幼蟲隨著齡期的增長過冷卻點逐漸升高,雌雄成蟲過冷卻點之間差異不顯著,但雌成蟲結冰點顯著低于雄成蟲。何莉梅等對草地貪夜蛾的發育起點溫度、有效積溫和發育歷期等指標測定結果顯示卵和1齡幼蟲的發育起點溫度較低,說明卵和1齡幼蟲對溫度的適應能力更強[14]。對于非滯育狀態的昆蟲,主觀印象認為其抗寒能力較低,本文的研究結果顯示卵和1齡幼蟲的過冷卻點(-25.45±0.25)℃、(-19.02±0.41)℃,比甜菜夜蛾卵和1齡幼蟲的過冷卻點(-14.60±1.80)℃、(-15.86±1.43)℃[13,15]和小地老虎的卵和1齡幼蟲過冷卻點-9.5℃和-8.8℃[9]明顯偏低。蛹的過冷卻點為(-17.23±0.27)℃比黏蟲的(-15.07±0.561)℃[16]、甜菜夜蛾(-15.55±1.13)℃[13,15]、勞氏黏蟲(-16.20±0.57)℃[17]偏低,比滯育狀態下的草地螟幼蟲(-25.82±0.26)℃、旋幽夜蛾蛹(-23.67±2.63)℃[1819]偏高。上述研究結果表明草地貪夜蛾可能比非滯育且遷飛的其他昆蟲如小地老虎、甜菜夜蛾、黏蟲等具有更強的耐寒能力。 本文對草地貪夜蛾的過冷卻點測定發現同一發育階段個體的過冷卻點有一定的變異,除卵以外,其余蟲態的過冷卻點頻次分布均符合正態分布,結冰點的頻次分布除卵、5齡幼蟲、6齡幼蟲、預蛹外,其余各蟲態冰點頻次均符合正態分布。頻次分布表明,同一發育階段的不同個體之間耐寒性存在差異。Klok和Chown認為這種差異現象是變溫動物的一種適應策略,在遭受突如其來的寒冷天氣時,抗逆性強的個體也能存活下來[20]。另外,影響昆蟲過冷卻點和結冰點測定結果的因素有很多,如不同地理種群、不同季節的蟲源、昆蟲本身的生長發育狀況、室內和野外種群、不同取食狀態以及不同測定方法等都會影響其測定結果[15]。根據對其他昆蟲的研究結果,草地貪夜蛾的抗寒力和越冬區的劃分,除測定其過冷卻點和結冰點外,還應結合潛在越冬區的實際蟲情普查、寄主調查、田間籠罩試驗及氣候條件等因素進行綜合分析,獲得該蟲在不同環境因子下發生為害規律等科學數據,以便今后更加全面系統地了解草地貪夜蛾的抗寒能力,為其越冬區劃分和預測預報等提供更全面可靠的數據支持。 參考文獻 [1]?姜玉英, 劉杰, 朱曉明. 草地貪夜蛾侵入我國的發生動態與未來趨勢分析[J]. 中國植保導刊, 2019, 39(2): 3335. [2]?張智, 張云慧, 姜玉英, 等. 草地貪夜蛾覆毛卵塊與絨繭蜂繭塊的識別特征[J/OL]. 植物保護: 14[20190823]. https:∥doi. org/10. 16688/j. zwbh. 2019402. [3]?全國農業技術推廣服務中心. 當前玉米重大病蟲害發生動態[EB/OL]. (20190823). https:∥www.natesc.org.cn/Html/2019-08-23/28092-151760-2019-08-23-459724.html. [4]?LEATHER S R, WALTERS K F A, BALE J S. The ecology of insect overwintering [M]. Cambridge: Cambridge University Press, 1993. [5]?RENAULT D,SALIN C,VANNIER G,et al.Survival at low temperatures in insects: What is the ecological significance of the supercooling point?[J].CryoLetters,2002(4),23:217228. [6]?景曉紅, 康樂. 昆蟲耐寒性的測定與評價方法[J]. 昆蟲知識, 2004, 40(1): 710. [7]?SALT R W. Principles of insect cold?hardiness [J]. Annual Review of Entomology, 1961, 6(3): 5574. [8]?陳若箎,趙健,徐秀媛.褐飛虱越冬溫度指標的研究[J].昆蟲學報,1982,25(4):390396. [9]?王蔭長,陳長琨,尤子平.小地老虎抗寒能力的研究[J].植物保護學報,1987,14(1):914. [10]吳孔明,郭予元.棉鈴蟲的抗寒能力[J].生態學報,1997,17(3):298302. [11]SPARKS A N. A review of the biology of the fall armyworm[J]. Florida Entomologist, 1979, 62(2): 8287. [12]BOTHA A S, ERASMUS A, DU PLESSIS H, et al. Efficacy of Bt maize for control of Spodoptera frugiperda (Lepidoptera: Noctuidae) in South Africa [J/OL]. Journal of Economic Entomology, 2019, doi: 10. 1093/jee/toz048. [13]江幸福. 甜菜夜蛾抗寒與越冬能力研究[J]. 生態學報, 2011, 21(10): 15751582. [14]何莉梅, 葛世帥, 陳玉超, 等, 草地貪夜蛾的發育起點溫度、有效積溫和發育歷期預測模型[J/OL]. 植物保護: 111[20190823]. https:∥doi. org/10. 16688/j. zwbh. 2019409. [15]韓蘭芝, 翟保平, 張孝羲. 甜菜夜蛾的抗寒力研究[J]. 植物保護學報, 2005, 32(2): 169173. [16]段云, 鞏中軍, 李慧玲, 等. 黏蟲幼蟲和蛹過冷卻點及結冰點的測定[J]. 植物保護, 2016, 42(4): 147150. [17]秦建洋, 蘭建軍, 張蕾, 等, 勞氏黏蟲幼蟲和蛹過冷卻點及結冰點的測定[J]. 中國植保導刊, 2018, 38(8): 3338. [18]李朝緒, 羅禮智, 潘賢麗. 草地螟滯育和非滯育幼蟲抗寒能力的研究[J]. 植物保護, 2006, 32(2): 4144. [19]趙琦, 張云慧, 韓二賓, 等. 旋幽夜蛾各蟲態的過冷卻點測定[J]. 植物保護, 2011, 37(2): 6366. [20]KLOK C J, CHOWN S L. Interactions between desiccation resistance, host plant contact and the thermal biology of aleaf?dwelling sub?antarctic caterpillar, Embryonopsis halticella (Lepidoptera: Yponomeutidae) [J]. Journal of Insect Physiology, 1998, 44: 615628. (責任編輯:楊明麗)
