假眼小綠葉蟬與其天敵的空間關系及最佳樣方確定
2019-02-10 10:50:51劉愛國張書平畢守東余燕李尚閆萍周夏芝鄒運鼎
植物保護 2019年6期
劉愛國 張書平 畢守東 余燕 李尚 閆萍 周夏芝 鄒運鼎


摘要 :為了分析在聚塊大小不同時天敵對假眼小綠葉蟬Empoasca vitis Gothe空間上跟隨關系的密切程度、聚集原因和聚集范圍,為評價假眼小綠葉蟬的天敵優勢種提供科學依據,用聚塊樣方方差分析法、灰色關聯度法、空間聚集強度指數、種群聚集均數法和ρ指數法對安徽省合肥市‘烏牛早和‘白毫早茶園不同大小聚塊條件下的假眼小綠葉蟬及其7種蜘蛛類的天敵空間關系進行分析。假眼小綠葉蟬與其7種蜘蛛類天敵均方差峰值時的聚塊樣方數的關聯度分析結果表明:‘烏牛早茶園中與假眼小綠葉蟬空間上跟隨關系密切的前四種天敵依次是茶色新圓蛛Neoscona theisi (0.753 5)、八斑球腹蛛Theridion octomaculatum (0.720 1)、錐腹肖蛸Tetragnatha maxillosa (0.681 3)和草間小黑蛛Erigonidium graminicolum (0.644 2)。‘白毫早茶園中與假眼小綠葉蟬空間上跟隨關系密切的前四種天敵依次是粽管巢蛛Clubiona japonicola (0.823 5)、鱗紋肖蛸Tetragnatha squamata (0.800 9)、錐腹肖蛸(0.794 2)和茶色新圓蛛(0.794 2)。兩種茶園前四種天敵中相同的天敵是茶色新圓蛛和錐腹肖蛸。假眼小綠葉蟬在聚塊內基本樣方數k為1、2、4、8時,隨著聚塊內基本樣方數的增多,聚集分布格局時的擴散系數C不斷增大,均勻和隨機格局時擴散系數不斷減小。聚塊內基本樣方數k為2、4、8時與k為1時的假眼小綠葉蟬的空間分布聚集程度差異均不顯著。假眼小綠葉蟬的種群聚集均數λ多數情況均大于2,其聚集是該蟲本身原因引起的,假眼小綠葉蟬在種群聚集均數λ為正值時,隨著聚塊內基本樣方數的增加,則種群聚集均數λ不斷增大。用假眼小綠葉蟬不同大小聚塊的ρ指數判斷個體群聚集時的最小范圍是聚塊中有1個基本樣方,即本文的4 m2。該研究為該蟲抽樣時確定樣方大小提供了科學依據。
關鍵詞 :假眼小綠葉蟬;?天敵;?空間關系;?聚塊樣方方差分析;?灰色關聯度分析
中圖分類號:
Q 968.1
文獻標識碼:?A
DOI:?10.16688/j.zwbh.2018456
Spatial relationships between Empoasca vitis and its natural
enemies and determination of optimal sample size
LIU Aiguo1,?ZHANG Shuping1,2,?BI Shoudong1,?YU Yan1,2,LI Shang1,2,?YAN Ping1,?ZHOU Xiazhi2,?ZOU Yunding2
(1. School of Science, Anhui Agricultural University, Hefei?230036, China;
2. School of Forestry and Landscape Architecture, Anhui Agricultural University, Hefei?230036, China)
Abstract
In order to analyze the closeness of the relationship between the natural enemy and Empoasca vitis Gothe, the reason and the range of agglomeration were analyzed. It helps provide a scientific basis for evaluating the dominant species of natural enemy of E.vitis. By means of block square variance analysis, grey correlation degree method, spatial aggregation intensity index, the population aggregation mean method and ρ index method, the spatial relationships between E.vitis and its natural enemies (seven spiders) were analyzed in ‘Wuniuzao and ‘Baihaozao tea gardens of Hefei, Anhui province. The correlation degree analysis of the number of blocks at the peak of the mean square deviation of 7 species of spiders and E.vitis showed that the first four natural enemies spatially closely related to E.vitis were Neoscona theisi (0.753 5), Theridion octomaculatum (0.720 1), Tetragnatha maxillosa (0.681 3) and Erigonidium graminicolum (0.644 2) in ‘Wuniuzao tea garden. In ‘Baihaozao tea garden, the first four natural enemies spatially closely related to E.vitis were Clubiona japonicola (0.823 5), Tetragnatha squamata (0.800 9), T. maxillosa (0.794 2) and Neoscona theisi (0.794 2). With the increase of the number of basic samples in the cluster, the diffusion coefficient C increased, and the diffusion coefficient decreased when the distribution pattern was uniform and random. There was no significant difference in the spatial aggregation degree of E.vitis and its natural enemies between k=2, 4, 8 and k=1. In most cases, the population aggregation mean λ was greater than 2, and its aggregation was caused by the insect itself. When the population aggregation mean λ was positive, the natural enemy and E.vitis increased with the increase of the basic sample size in the agglomeration. The aggregation mean λ of the population increased continuously. The minimum range of ρ index required one basic sample in the cluster. For example, it was 4 m2 in this study. It provides a scientific basis for determining the sample size when the insect is sampled.
Key words
Empoasca vitis;?natural enemy;?spatial relationship;?analysis of square variance of cluster sample; grey correlation analysis
茶樹害蟲是影響茶葉產量和品質的主要因子之一。自然天敵是持續影響茶樹害蟲種群消長的重要生態因子。假眼小綠葉蟬Empoasca vitis是重要的茶樹害蟲[1],其發生規律及其天敵種類有大量研究報道。Chen等[23]報道了茶樹害蟲天敵蜘蛛三者之間的化學聯系及茶樹的抗性機制。研究者研究了次生物質對害蟲和天敵行為的影響[46]。王沅江等[7]、扈克明等[8]報道瓢蟲和蜘蛛都取食假眼小綠葉蟬。曾莉等[9]研究了茶樹品種對假眼小綠葉蟬的抗性。鈕羽群等[10]研究了不同迷迭香揮發物組合對假眼小綠葉蟬行為的調控。
黎健龍等[11]研究了周邊不同生境條件對茶園蜘蛛群落及葉蟬時空結構的影響。黨鳳花等[12]報道了江淮地區‘龍井43茶園假眼小綠葉蟬的主要天敵優勢種,王振興等[13]、柯勝兵[14]、畢守東等[15]報道了江淮地區和大別山區不同海拔茶園假眼小綠葉蟬及其天敵的分布與差異。周夏芝等[16]研究了大別山區低海拔的天柱山群體種茶園中天敵對假眼小綠葉蟬的空間跟隨關系。劉飛飛等[17]探討了江淮地區‘黃山大葉種茶園春夏季與秋冬季假眼小綠葉蟬天敵優勢種的差異。方國飛等[18]研究了合肥地區‘平陽特早茶園不同年度間和季節間假眼小綠葉蟬與其天敵在數量、時間和空間上關系的差異。劉飛飛等[19]研究了大別山區不同種類蜘蛛天敵與假眼小綠葉蟬空間跟隨關系的差異。韓寶瑜等[20]報道了間作密植和單作茶園對假眼小綠葉蟬等害蟲及其天敵種群數量的影響及群落多樣性和穩定性的差異。謝振倫[21]研究了茶園3種蜘蛛對假眼小綠葉蟬的捕食量。抽樣調查的樣方大小和樣方數量多少不僅與成本有關,而且直接影響調查結果的準確性,Morris 1995年對樣方提出了8條標準,其中兩條都涉及樣方大小[22]。假眼小綠葉蟬在該方面的研究未見報道。本文采用聚塊樣方方差分析法等研究聚塊大小對假眼小綠葉蟬及其天敵聚塊數量均方差的影響,以及假眼小綠葉蟬個體群聚集的最小范圍,為評定假眼小綠葉蟬天敵優勢種及確定最佳抽樣樣方大小提供科學依據。
1?材料與方法
1.1?調查地點和時間
調查地點為安徽農業大學科技示范園茶園,調查茶樹品種為樹齡13年的‘烏牛早和‘白毫早,茶園面積為0.2 hm2。根據合肥地區茶園害蟲及天敵的發生特點[19],即3月下旬才有少量種類害蟲發生,所以第一次取樣時間選在3月下旬。調查時間為2015年3月28日—11月14日。2016年3月27日—11月17日,約15 d調查1次,2015年共調查17次,2016年調查了16次。茶園周邊為其他品種茶園,茶園按常規措施管理,但不施用農藥。
1.2?調查方法
采用平行跳躍法隨機在各個茶園選取3行,茶樹行寬為2 m,每行間隔1 m取2 m長的樣方;即1個樣方的面積為4 m2。每行茶樹選取10個樣方,共取30個樣方。先目測調查,每樣方隨機選取10片葉,調查一些不易振落害蟲及天敵種類和個體數,然后用沾有洗衣粉水液的搪瓷盤對樣方中的所有枝條進行盤拍(搪瓷盤口長為40 cm,寬30 cm,洗衣粉水溶液濃度為1 000倍),調查記載害蟲及其天敵物種數和個體數,對于不能準確鑒定的物種樣本編號保存,裝瓶帶回室內鑒定。
1.3?數學分析方法
根據聚塊樣方方差分析的要求,參與數學分析的資料是第1個樣方連續至第24個樣方的調查資料。
1.3.1?假眼小綠葉蟬空間動態的聚塊樣方方差分析
聚塊樣方方差分析是在不同大小樣方上的方差分析,是一種簡單、有效的生態學空間格局分析方法。該法要求供試田塊上的樣方在空間上相互連接,隨著聚塊所包含的基本樣方數k從1,2,4,8…(指數級數)不斷增加,聚塊方差值常隨之改變,通過不同大小聚塊方差值的變化,了解研究對象隨尺度增大的變化動態[23]。
在一樣帶上連續分布的樣方,以假眼小綠葉蟬或天敵在每個聚塊上的數量為變量(X),讓聚塊內基本樣方數成指數增大,計算其均方差MS。均方差的計算公式為:MS(k)=1n∑n-2k+1i=1(Xi-Xi+1)2,其中i為每次新生成聚塊序列數,k為聚塊內所含基本樣方數,n為參與分析的總樣方數,本文取24,即k的最大可能值為12,但由于k按指數級增加,因而取不到12,所以本文聚塊內基本樣方數為1,2,4,8。聚塊樣方方差分析的目的是研究聚塊大小對方差的影響,如果均方差在某一聚塊大小上出現峰值,則表明田間害蟲空間分布具有規律性。
1.3.2?假眼小綠葉蟬與7種天敵在均方差峰值時基本樣方數上的灰色關聯度分析
將假眼小綠葉蟬及其7種天敵均方差峰值時的基本樣方數(即聚塊空間大小)分別看作一個本征系統,假眼小綠葉蟬每次峰值時的基本樣方數作為該系統的參照序列,其各種天敵的基本樣方數作為該系統的比較序列,不同時間點上的假眼小綠葉蟬聚集空間大小及其天敵的聚塊基本樣方數作為序列在第H個樣方上的效果白化值,進行雙序列關系分析,求兩者間的關聯度,關聯度大小反映了天敵對假眼小綠葉蟬空間上跟隨關系的密切程度,關聯度值越大,表明兩者在空間上關系越密切[24]。
1.3.3?假眼小綠葉蟬空間格局及其差異原因分析
本文采用Poisson擴散系數C分析測定假眼小綠葉蟬的空間分布格局[25]。用David等提出的|w|公式,|w|=-12ln(S21/x-1S22/x-2),判斷k為2、4、8時與1時的聚集程度差異,S21,S22,x-1,x-2分別為聚塊內基本樣方數為2,4,8與為1時的兩種群的方差和均數,若|w|>2.5/n-1,則按5%水平認為兩者的空間分布格局顯著不同[26]。用Arbous等[27]提出的種群聚集均數(λ)公式,λ=x-2k·v,分析不同聚塊大小條件下假眼小綠葉蟬的聚集原因,式中k=x-2/(s2-x-),S2為方差,v為自由度等于2k時的χ20.05值。用Blackith提出λ值判斷標準,分析引起種群聚集的原因[28]。用Iwao[29]的ρ指數公式,ρ=i-i-1x-i-x-i-1,依據一系列不同大小聚塊的ρ指數來評定假眼小綠葉蟬個體群聚集時占據的最小空間,即最小的樣方面積。式中i和i-1為i和i-1聚塊的平均擁擠度,x-i和x-i-1為i和i-1聚塊的平均密度。
2?結果與分析
2015年調查結果表明,‘烏牛早茶園共采集到假眼小綠葉蟬3 227頭,37種捕食性天敵,共7 374頭。2016年茶園共采集到假眼小綠葉蟬5 046頭,捕食性天敵40種,共5 002頭。2015年‘白毫早茶園共采集到假眼小綠葉蟬1 896頭,42種捕食性天敵,共6 111頭。2016年茶園共采集到假眼小綠葉蟬1 094頭,捕食性天敵31種,共6 045頭。蜘蛛類天敵是假眼小綠葉蟬的主要捕食性天敵,2015年調查數量大于225頭,2016年調查數量大于144頭的前7種蜘蛛是鱗紋肖蛸Tetragnatha squamata,錐腹肖蛸Tetragnatha maxillosa,草間小黑蛛Erigonidium graminicolum,三突花蟹蛛Misumenops tricuspidatus,八斑球腹蛛Theridion octomaculatum,粽管巢蛛Clubiona japonicola和茶色新圓蛛Neoscona theisi。兩種茶園2015年7種蜘蛛數量共7 937頭,占全年捕食性天敵的58.86%,2016年7種蜘蛛數量為8 194頭,占全年捕食性天敵的74.17%,本文將7種蜘蛛作為假眼小綠葉蟬的主要天敵。
2.1?假眼小綠葉蟬及其7種天敵空間關系的聚塊樣方方差分析
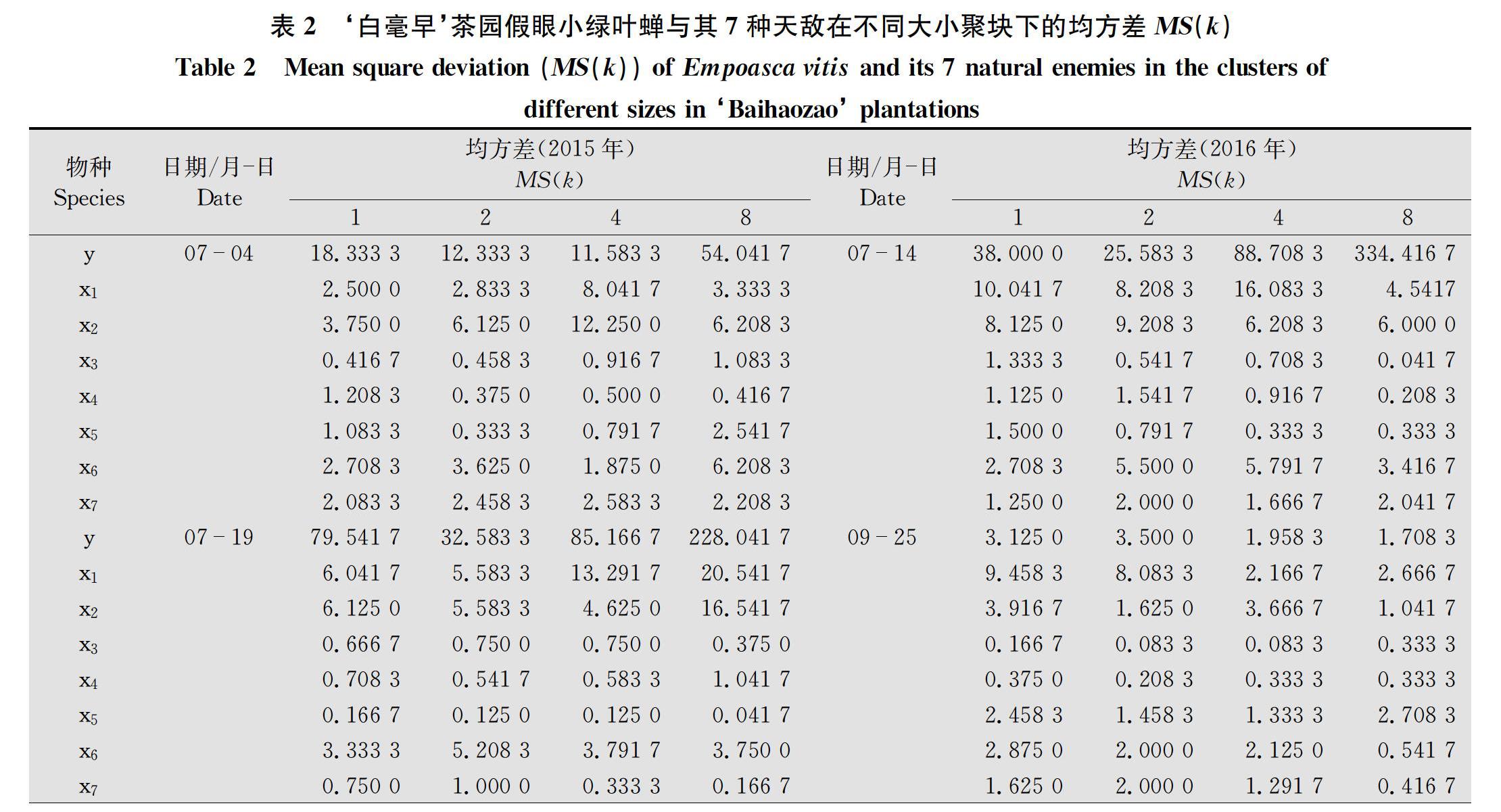
為了體現研究結果的代表性,假眼小綠葉蟬選擇數量多的、數量中等的和數量少的3種類型作為研究材料。‘烏牛早茶園2015年選擇5月10日(47頭)、7月19日(548頭)、9月13日(358頭),2016年選擇5月12日(49頭)、7月14日(386頭)、11月2日(186頭);‘白毫早茶園2015年選擇7月4日(275頭)、7月19日(256頭)、9月13日(133頭),2016年選擇7月14日(313頭)、9月25日(38頭)、11月2日(148頭)的假眼小綠葉蟬及其天敵作為研究對象,將假眼小綠葉蟬及其7種天敵的聚塊樣方方差分析的均方差列于表1和表2。
1) y:假眼小綠葉蟬;x1:鱗紋肖蛸;x2:錐腹肖蛸;x3:草間小黑蛛;x4:三突花蟹蛛;x5:八斑球腹蛛;x6:粽管巢蛛;x7:茶色新圓蛛。下同。y: Empoasca vitis; x1:Tetragnatha squamata; x2:Tetragnatha maxillosa; x3:Erigonidium graminicolum; x4:Misumenops tricuspidatus; x5:Theridion octomaculatum; x6:Clubiona japonicola; x7:Neoscona theisi. The same below.
從表中可看出,‘烏牛早2015年5月10日、7月19日、9月13日、2016年7月14日、11月2日和‘白毫早2015年9月13日、2016年9月25日的假眼小綠葉蟬的均方差均有一個峰值,‘烏牛早2016年5月12日和‘白毫早2015年7月4日、7月19日、2016年7月14日有兩個高峰。7種蜘蛛的均方差均有1~2個峰值,表明假眼小綠葉蟬和7種天敵的空間分布均具有一定的規律性,可對其關系進行深入研究。為了分析7種天敵與假眼小綠葉蟬空間關系密切程度,將7種蜘蛛與假眼小綠葉蟬均方差峰值時聚塊內基本樣方數列于表3,并進行灰色關聯度分析,結果表明,‘烏牛早茶園茶色新圓蛛與假眼小綠葉蟬的關聯度最大,為0.753 5,其次是八斑球腹蛛,為0.720 1,第三是錐腹肖蛸,為0.681 3,鱗紋肖蛸、草間小黑蛛、三突花蟹蛛和粽管巢蛛與假眼小綠葉蟬的關聯度分別是0.589 6、0.644 2、0.596 5和0.595 7。表明茶色新圓蛛、八斑球腹蛛和錐腹肖蛸與假眼小綠葉蟬空間關系密切。‘白毫早茶園粽管巢蛛與假眼小綠葉蟬的關聯度最大,關聯度值為0.823 5,其次是鱗紋肖蛸(0.800 9),第三是錐腹肖蛸(0.794 2)和茶色新圓蛛(0.794 2),草間小黑蛛、三突花蟹蛛和八斑球腹蛛與假眼小綠葉蟬的關聯度分別是0.625 4、0.672 2和0.621 5,兩品種茶園中前4位中相同的天敵是茶色新圓蛛和錐腹肖蛸。
2.2?聚塊大小對假眼小綠葉蟬空間分布格局的影響
用擴散系數C值作為判斷空間分布格局類型的依據,將假眼小綠葉蟬在不同聚塊大小條件下的擴散系數C值列于表4。可看出,‘烏牛早茶園假眼小綠葉蟬2015年7月19日、9月13日和2016年7月14日聚塊樣方數從1到8的C值均大于F0.05,表明是聚集分布格局,這3個時間的假眼小綠葉蟬依次是548頭、358頭和386頭,即該蟲數量較多時是聚集格局,而2015年5月10日假眼小綠葉蟬為47頭,2016年5月12日假眼小綠葉蟬為49頭,2016年11月2日假眼小綠葉蟬為186頭,這些都不是聚集格局,似乎表明聚集格局與一定的蟲口數量有關。但同一個時間與基本樣方數k為1和k為2、4、8時相比,如2015年5月10日,個體數是47頭,k為2、4、8時是聚集格局,只有k為1時不是聚集格局,k為2、4、8時的標準差為2.429 3、4.070 2和6.506 4,而k為1時只有1.768 8,表明決定分布格局的原因與標準差大小有關,即與蟲口數量的離散性大小有關。同時可看出,這兩個時段,隨著聚塊內樣方數的增加,C值不斷增大。假眼小綠葉蟬在均勻分布和隨機分布時,C值變化的總趨勢是,隨著聚塊內基本樣方數的增加,C值不斷變小。
1) C值的判別標準:n1=23,n2=∞,F0.05=1.51;n1=11,n2=∞,F0.05=1.79;n1=5,n2=∞,F0.05=2.21;n1=2,n2=∞,F0.05=3。Criteria for discriminating C values: n1=23,n2=∞,F0.05=1.51;n1=11,n2=∞,F0.05=1.79;n1=5,n2=∞,F0.05=2.21;n1=2,n2=∞,F0.05=3.
2.3?假眼小綠葉蟬在聚塊內基本樣方數為2、4、8時與為1時的聚集程度差異
用David和Moore提出的比較總體聚集程度的方法,求出|W|值,結果列于表5。可看出假眼小綠葉蟬在聚塊內基本樣方數為2、4、8時與為1時的|W|值均小于|W|0.05,表明聚塊大小對假眼小綠葉蟬空間聚集程度影響不顯著。
2.4?假眼小綠葉蟬在不同聚塊大小條件下種群聚集均數的變化及其聚集原因
將假眼小綠葉蟬在不同聚塊大小條件下的種群聚集均數λ列于表6,假眼小綠葉蟬兩年的6個時間點不同聚塊大小條件下的λ值,除了‘烏牛早茶園2015年5月10日k為1時和2016年5月12日k為1時的λ值小于2,其余的λ值均大于2,且隨著聚塊內基本樣方數的增加,λ值不斷增大,即隨著聚塊面積增大,λ值不斷變大。當λ值為負值時,隨著聚塊內基本樣方數的增加,λ值不斷變小。由表6可看出,假眼小綠葉蟬在兩種茶園兩年12次中有8次的λ值均大于2,表明其聚集是由該害蟲本身原因引起的。
2.5?不同聚塊大小條件下假眼小綠葉蟬的ρ指數
表7為不同聚塊條件下假眼小綠葉蟬的ρ指數,以茶園為標準可看出兩年‘烏牛早茶園聚塊內基本樣方數k由2到1時,平均ρ指數最大,為1.142 7±0.059 6;兩年‘白毫早茶園聚塊內基本樣方數k由8到4時,平均ρ指數最大,為1.066 6±0.025 6。以年為標準兩品種合并來看,2015年聚塊內基本樣方數k由4到2時,平均ρ指數最大,為1.125 7±0.047 4,2016年聚塊內基本樣方數k由2到1時,平均ρ指數最大,為1.081 9±0.01,以年和茶園為標準,聚塊基本樣方數由2到1的ρ指數平均值最大4次中出現兩次,由4到2和8到4各出現一次,因此,綜合考慮,假眼小綠葉蟬個體群在聚集格局時種群的最小面積是聚塊內基本樣方數為1個,按照本文的調查設計,即每一聚塊面積是4 m2,即取樣調查假眼小綠葉蟬時,每個樣方應不小于4 m2。
1) 假眼小綠葉蟬的數量(N):‘烏牛早茶園2015年5月10日為47頭,7月19日為548頭,9月13日為358頭;2016年5月12日為49頭,7月14日為386頭,11月2日為186頭;‘白毫早茶園2015年7月4日為275頭,7月19日為256頭,9月13日為133頭;2016年7月14日為313頭,9月25日為38頭,11月2日為148頭。
The number of Empoasca vitis in ‘Wuniuzao tea garden: 47 individuals on May 10, 548 individuals on July 19 and 358 individuals on September 13, 2015; 49 individuals on May 12, 386 individuals on July 14 and 186 individuals on November 2, 2016; The number of Empoasca vitis in ‘Baihaozao tea garden: 275 individuals on July 4, 256 individuals on July 19 and 133 individuals on September 13, 2015; 313 individuals on July 14, 38 individuals on September 25 and 148 individuals on November 2, 2016.
3?小結與討論
運用聚塊樣方方差分析法、灰色關聯度法和聚集強度指數法相結合研究了兩種茶園7種蜘蛛類天敵對假眼小綠葉蟬在空間上的跟隨關系及聚集原因等。假眼小綠葉蟬與其7種蜘蛛類天敵均方差峰值時的聚塊大小(聚塊內的基本樣方數)的灰色關聯度分析及擴散系數、種群聚集均數和ρ指數的分析結果表明:1)‘烏牛早茶園與假眼小綠葉蟬空間上跟隨關系密切的前四位天敵是茶色新圓蛛、八斑球腹蛛、錐腹肖蛸和草間小黑蛛;‘白毫早茶園與假眼小綠葉蟬空間上跟隨關系密切的前四位天敵是粽管巢蛛、鱗紋肖蛸、錐腹肖蛸和茶色新圓蛛。2)假眼小綠葉蟬在聚塊內基本樣方數k為1~8時,隨著聚塊內基本樣方數k的增加,聚集格局時的擴散系數C值一直增大,而均勻格局和隨機格局時C值不斷變小:3)假眼小綠葉蟬在不同大小聚塊條件下的聚集多是由該蟲本身原因引起的;4)假眼小綠葉蟬在種群聚集均數λ為正值時,隨著聚塊內基本樣方數k的增加,λ值不斷變大;5)用不同大小聚塊的ρ指數判斷假眼小綠葉蟬種群聚集時的最小面積為聚塊內有1個基本樣方,即4 m2,為該蟲抽樣時確定樣方大小提供了科學依據。
周夏芝等[16]、王振興等[12]運用空間生態位重疊指數和相似性系數大小來判斷與假眼小綠葉蟬空間跟隨密切的天敵,其結果均分別有八斑球腹蛛、錐腹肖蛸或茶色新圓蛛,與本文用聚塊樣方方差分析的結果一致。劉飛飛等[30]和王建盼等[31]分別用聚塊樣方方差分析法與其他方法相結合研究了大別山區天柱山群體種茶園蜘蛛類天敵與茶尺蠖Ectropis obliqua幼蟲的空間關系及柑橘粉虱Dialeurodes citri Ashmead與其捕食天敵之間的空間關系,在聚集格局、均勻格局和隨機格局時擴散系數C和λ值變化動態上的研究結論與本文一致。
參考文獻
[1]?張漢鵠, 譚濟才. 中國茶樹害蟲及其無公害治理[M]. 合肥: 安徽科學技術出版社, 2004.
[2]?CHEN Zongmao, XU Ning, HAN Baoyu. Role of volatile allelochemicals on host location of tea pests and host foraging of natural enemies in tea ecosystem [C]. Scientific Program and Abstracts of the First Asua?Pacific Conference on Chemical Ecology, 1999: 8183.
[3]?CHEN Zongmao, SUN Xiaoling, DONG Wenxia. Genetics and chemistry of the resistance of tea plant to pests [M]. Global Tea Breeding, 2012: 343360.
[4]?PERSONS M H, RYPSTRA A L. Preference for chemical cues associated with recent prey in the wolf spider Hogna helluo (Araneae: Lycosidae) [J]. Ethology, 2000, 106(1): 2735.
[5]?CLARK R J, JACKSON R R, CUTLER B. Chemical cues from ants influence predatory behavior in Habrocestum pulex, an ant?eating jumping spider (Araneae, Salticidae) [J].Journal of Arachnology, 2000, 28(3): 309318.
[6]?XIAO Yonghong, ZHANG Jiaxu, LI Shuqiang. A two?component female produced pheromone of the spider Pholcus beijingensis [J]. Journal of Chemical Ecology, 2009, 35(7): 769778.
[7]?王沅江, 謝振倫, 龐雄飛. 假眼小綠葉蟬及天敵蜘蛛生態位的研究[J]. 茶葉科學, 2008, 28(6): 401406.
[8]?扈克明, 張艷梅, 王佳芳, 等. 不同茶樹品種間小綠葉蟬類群數量動態與抗蟲性比較[J]. 茶葉科學, 2003, 23(1): 5760.
[9]?曾莉, 王平盛, 許玫. 茶樹對假眼小綠葉蟬的抗性研究[J]. 茶葉科學, 2001(2): 9093.
[10]鈕羽群, 王夢馨, 崔林, 等. 迷迭香揮發物不同組合對假眼小綠葉蟬行為的調控[J]. 生態學報, 2015, 35(7): 23802387.
[11]黎健龍,唐勁馳,黎秀娣,等.周邊不同生境條件對茶園蜘蛛群落及葉蟬種群時空結構的影響[J].生態學報,2014,34(19): 22162227.
[12]黨鳳花, 公茂蓮, 畢守東, 等. 茶園3種天敵主要害蟲與其多種天敵之間密切程度的比較[J]. 浙江大學學報(農業與生命科學版), 2010, 36(5): 513520.
[13]王振興,李尚,王建盼,等.兩品種茶園茶蚜和假眼小綠葉蟬天敵優勢種比較[J].安徽農業大學學報,2016,43(3):350358.
[14]柯勝兵, 黨鳳花, 畢守東, 等. 不同海拔茶園害蟲、天敵種群及其群落結構差異[J]. 生態學報, 2011, 31(14): 41614168.
[15]畢守東, 柯勝兵, 徐勁峰, 等. 3種海拔高度茶園中2種害蟲與其天敵間的數量和空間關系[J]. 生態學報, 2011, 31(2): 455464.
[16]周夏芝, 畢守東, 黃勃, 等. 茶園主要天敵對4種害蟲的空間跟隨關系[J]. 華南農業大學學報, 2013, 3(4): 489498.
[17]劉飛飛, 王建盼, 林源, 等. 不同季節黃山大葉種茶園主要害蟲的捕食性天敵優勢種比較[J]. 華南農業大學學報, 2014, 35(6): 6773.
[18]方國飛, 畢守東, 劉飛飛, 等. 平陽特早茶園3種主要害蟲與其天敵在數量、時間和空間上的關系[J]. 安徽農業大學學報, 2015, 42(6): 860867.
[19]劉飛飛,柯勝兵,王建盼,等.假眼小綠葉蟬與捕食性天敵蜘蛛的空間關系研究[J].西北農林科技大學學報(自然科學版),2016,44(6): 99110.
[20]韓寶瑜, 江昌俊, 李卓民. 間作密植和單行茶園節肢動物群落組成差異[J]. 生態學報, 2001, 21(4): 646652.
[21]謝振倫. 茶園三種蜘蛛對假眼小綠葉蟬捕食量的觀察[J]. 廣東茶葉, 1996(2): 3234.
[22]MORRIS R F. The development of sampling techniques for forest insect defoliators, with particular reference to the spruce budworm [J].Canadian Journal of Zoology,1995,33:225294.
[23]傅伯杰, 陳利頂, 馬克明, 等. 景觀生態原理及應用[M]. 北京: 科學出版社, 2003: 216217.
[24]鄧聚龍. 灰色系統理論教程[M]. 武漢: 華中科技大學出版社, 1990: 3384.
[25]鄒運鼎, 王弘法. 農林昆蟲生態學[M]. 合肥: 安徽科學技術出版社, 1989: 152157.
[26]DAVID F N, MOORE P G. Notes on contagious distribution in plant population [J]. Annals of Botany, 1954, 18: 4753.
[27]ARBOUS A G, KERRICH J E.Accident statistics and the concept of accident?proneness [J]. Biometrics, 1951, 7(4): 340432.
[28]BLACKITH R E.Nearest?neighbour distance measurements for the estimation of animal populations [J]. Ecology, 1958, 39: 147150.
[29]IWAO S. Application of the m*?m method to the analysis of spatial patterns by changing the quadrat size [J].Researches on Population Ecology, 1972, 14(1): 97128.
[30]劉飛飛, 柯勝兵, 王建盼, 等. 蜘蛛類天敵與茶尺蠖幼蟲空間關系的聚塊樣方方差分析[J]. 浙江大學學報(農業與生命科學版), 2015, 41(2): 133146.
[31]王建盼,覃盛,劉飛飛,等.柑橘粉虱與主要捕食性天敵之間的空間關系研究[J].中國生態農業學報,2015,23(4):454464.
(責任編輯:楊明麗)
