肺炎克雷伯氏菌漆酶基因的克隆及其在畢赤酵母中的高效表達
2019-02-23 05:25:22王建玲張妤彤路福平劉逸寒
天津科技大學學報 2019年1期
王建玲,桂 爽,付 禹,張妤彤,鄭 東,路福平,劉逸寒
(工業發酵微生物教育部重點實驗室,天津市工業微生物重點實驗室,工業酶國家工程實驗室,天津科技大學生物工程學院,天津 300457)
漆酶(laccase,EC 1.10.3.2),又稱漆酚氧化酶、多銅氧化酶,是一種含銅的多酚氧化酶,屬于銅藍氧化酶蛋白家族的一員[1–2].漆酶的作用底物較為廣泛,能夠催化包括多酚類、二胺、芳胺類、羧酸類在內的250多種有機物發生氧化、聚合等反應.由于漆酶具有特殊的催化性能和寬泛的作用底物,因此其應用較為廣泛,包括紡織業中的染料脫色、造紙工業中的生物漂白、食品行業中食品風味改良、檢測酚類污染物生物傳感器的構建和生物電子研發等[3–6].部分上述工業過程需要在高溫、強酸、強堿等惡劣條件下進行,因此,需要漆酶具有較好的溫度和pH穩定性[7].
漆酶主要存在于植物、真菌、少數昆蟲和細菌中,其中真菌漆酶來源最為廣泛,研究較為深入的真菌漆酶主要來自白腐真菌(C.thermophilium)[8]、射脈菌(Phlebia radiata)[9]、云芝栓孔菌(Trametes versicolor)[10]、鮮紅密孔菌(Pycnoporus cinnabarinus)[11]、糙皮側耳(P.ostreatus)[12]、毛木耳(Auricularia polytricha)[13]和黑木耳(Auricularia auricula)[14]等.目前,關于真菌漆酶的研究眾多,但其存在堿性條件下活性較低、熱穩定性較差和絲狀真菌生長周期長等缺點.細菌漆酶來源較少,目前已在生脂固氮螺菌(Azospirillum lipoferum)[15]、鏈霉菌(Streptomycetes)[16]、球形芽胞桿菌(Bacillus sphaericus)[17]、枯草芽胞桿菌(Bacillus subtilis)[18]、嗜熱棲熱菌(Thermus thermophilus)[19]及肺炎克雷伯氏菌(Klebsiella pneumoniae)[20]等細菌中發現過漆酶活性.與真菌來源漆酶相比,細菌漆酶在堿性條件下具有較好的催化活性和穩定性及存在 Cu2+抗性等優點[21–23].以 2,6-DMP 為底物時,細菌漆酶的最適溫度一般在 60~65℃,最適 pH 一般為 7.0~8.0,在 25~60℃和 pH 6.0~9.0范圍內穩定性較好[24–27].另外,黃俊等[20]篩選獲得一株產漆酶菌 Klebsiella sp.601,以 2,6-DMP為底物時,該漆酶在 pH 5.0~8.0條件下較穩定,在 pH 3.0~4.0、pH 9.0~10.0條件下,幾乎沒有殘余酶活力.本實驗室前期從中國西雙版納土壤中篩選獲得一株含漆酶基因(lac)的肺炎克雷伯氏菌[28],lac基因與上述Klebsiella sp.601來源漆酶基因相比,共有18個不同氨基酸,該漆酶以2,6-DMP 為底物時,在pH 5.0~9.0條件下較穩定,在 pH 3.0~4.0、pH 10.0~11.0條件下,其殘余酶活力在 20%以上,在 30~70℃條件下較穩定,其殘余酶活力在40%以上,表明該漆酶具有良好的 pH和溫度穩定性,但由于是在大腸桿菌中表達,難以實現高效合成.
本研究旨在通過構建前期篩選獲得的具有良好酶學特性的 lac畢赤酵母重組菌株,實現該重組漆酶的高效分泌表達,為其應用奠定基礎.
1 材料與方法
1.1 材料
1.1.1 菌種與質粒
肺炎克雷伯氏菌(Klebsiella pneumoniae)TCCC111142、質粒 pUC57-lacm(根據畢赤酵母的密碼子偏愛性優化的漆酶基因lacm克隆入pUC57中,由北京六合華大基因科技有限公司合成)、大腸桿菌(Escherichia coli)DH5α、畢赤酵母(Pichia pastoris)GS115、質粒pPIC9K均為本實驗室保存.
1.1.2 主要試劑與工具酶
Taq DNA 聚合酶、限制性內切酶 EcoRⅠ、NotⅠ,Takara公司;質粒快速提取試劑盒和 DNA 純化回收試劑盒,Omega公司;蛋白胨和酵母浸粉,Oxoid公司;其他試劑均為國產分析純.
1.1.3 培養基
LB固體培養基(g/L):酵母提取物5.0,胰蛋白胨10.0,NaCl 10.0,瓊脂 15.
YPD 固體培養基(g/L):酵母提取物 10,蛋白胨20,葡萄糖 20,瓊脂 20.
BMGY 液體培養基(g/L):酵母提取物 10,蛋白胨 20,YNB 13.4,甘油 5,生物素 0.04;1mol/L 磷酸鹽緩沖液(pH 6.0)100mL.
BMMY 液體培養基(g/L):酵母提取物 10,蛋白胨20,YNB 13.4,加入生物素0.04;1mol/L磷酸鹽緩沖液(pH 6.0)100mL,每日補加甲醇5mL.
MD 培養基(g/L):葡萄糖 20,瓊脂 15,YNB 13.4,生物素 0.04.
1.2 方法
1.2.1 肺炎克雷伯氏菌漆酶基因擴增
通過NCBI基因庫查找,根據已報道的肺炎克雷伯氏菌漆酶成熟肽基因,分析其保守序列,設計TCCC111142成熟肽編碼基因的擴增引物,上游引物A-3′(下劃線部分為加入的 EcoRⅠ酶切位點),下游劃線部分為加入的 NotⅠ酶切位點,雙下劃線部分為添加的6個組氨酸標簽,方便純化蛋白).
以肺炎克雷伯氏菌基因組為模板,以 P1和 P2為引物,進行 PCR擴增.擴增條件為:95℃預變性10min;95℃變性 30s,55℃退火 45s,72℃延伸1min 40s,30個循環;72℃延伸 10min.
將 PCR擴增產物進行測序后,獲得 lac基因序列,根據畢赤酵母的密碼子偏愛性優化該基因,由北京六合華大基因科技有限公司進行合成,得到新型漆酶基因lacm.
1.2.2 重組質粒的構建
重組質粒pUC57-lacm和質粒pPIC9K經EcoRⅠ和NotⅠ雙酶切并純化回收后,用T4 DNA連接酶連接,轉化大腸桿菌 DH5α感受態細胞中,經氨芐青霉素(Amp)抗性篩選,挑選陽性轉化子;提取質粒,并進行酶切驗證.
1.2.3 電轉化畢赤酵母
提取重組菌質粒 pPIC9K-lacm,用 SacⅠ酶切線性化并進行純化回收;制備畢赤酵母 GS115的感受態細胞;然后采用電轉化法將線性化的重組質粒pPIC9K-lacm轉化到畢赤酵母GS115,將電轉液涂布于MD平板,30℃靜置培養至轉化子出現.
1.2.4 酵母轉化子PCR鑒定
從 MD平板上挑選轉化子提取基因組,以其基因組為模板,用上述lacm基因特異引物進行PCR反應,凝膠電泳檢測 lacm基因是否插入到畢赤酵母GS115的染色體上.
1.2.5 漆酶在畢赤酵母中的誘導表達
選擇 PCR鑒定為陽性的菌落 GS115/pPIC9K-lacm和轉有空質粒的對照菌株 GS115/pPIC9K接種至 YPD 液體培養基中,30℃、220r/min培養 24h.以 3%的接種量轉接到新鮮 BMGY培養基中,繼續以 30℃、220r/min培養 24h,然后以 6000r/min離心 10min收集菌體,將菌體轉接到 BMMY培養基中.再以 30℃、220r/min培養,并每隔 12h補加一次甲醇,使其終體積分數保持在0.5%,培養120h后可得到重組漆酶(rLACM)的粗酶液.
1.2.6 純化rLACM
采用鎳柱純化親和層析法純化 rLACM,步驟如下:將含有 Ni樹脂裝入合適的純化柱中,樹脂體積為 2mL;用預冷的 Lysis Buffer緩沖溶液平衡樹脂2~5個床體積;將平衡后的樹脂與經過膜(0.22μm)過濾的粗酶液混合,在4℃條件下通過磁力攪拌器使二者結合 40~60min;將樹脂與粗酶液的結合液過柱;加入預冷的 Wash Buffer緩沖溶液 20mL,進行清洗雜蛋白;用500mmol/L咪唑的Elution Buffer緩沖溶液5mL進行洗脫,用于SDS-PAGE檢測目標蛋白的相對分子質量和純度.
1.2.7 表達產物的SDS-PAGE分析
重組菌株 GS115/pPIC9K-lacm 和對照菌GS115/pPIC9K 在30℃誘導培養結束后,取1mL發酵液離心去除菌體,取上清液和純化后 rLACM 各40μL進行SDS-PAGE分析.
1.2.8 漆酶活力測定及酶學性質考察
取 200μL的含 4mmol/L Cu2+的 0.1mol/L的檸檬酸–磷酸氫二鈉緩沖液(pH 2~8)或者含 4mmol/L Cu2+的 0.05mol/L的甘氨酸–氫氧化鈉緩沖液(pH 8.5~11),于96孔板中,70℃保溫1min.加入10μL的稀釋適當倍數的純酶液,混勻,70℃保溫1min.再加入30μL的底物(8mmol/L的2,6-DMP),混勻,記錄反應初始吸光度和反應3min時吸光度.在一定的條件下,每分鐘氧化1μmol的2,6-DMP所需要的酶量定義為一個酶活力單位(U).
用pH 8.0的磷酸緩沖溶液配制漆酶的2,6-DMP底物,分別在20~100℃測定其酶活力,確定rLACM的最適溫度.將酶液置于 pH 8.0緩沖液,在 30~80℃溫浴,每隔 1h取樣,在最適反應條件下測定其酶活力,考察其溫度穩定性.
分別用pH為2.0~11.0的磷酸緩沖溶液配制漆酶底物 2,6-DMP,在 70℃反應下,確定 rLACM 的最適pH.將酶液置于pH 3.0~10.0緩沖液,于70℃保溫35h,每隔5h測定rLACM酶活力,考察其pH穩定性.
2 結果與分析
2.1 重組質粒pPIC9K-lacm的構建
以肺炎克雷伯氏細菌的基因組為模板,以引物P1和P2擴增目的基因lac.在lac基因序列基礎上,基于畢赤酵母密碼子偏愛性,優化合成獲得 lacm基因,與 lac相比,共改變 406個堿基,GC含量由61.3%變為 48.4%.重組質粒 pUC57-lacm與質粒pPIC9K均經 EcoRⅠ和 NotⅠ雙酶切后連接,轉化大腸桿菌 DH5α感受態細胞中,經 Amp抗性篩選,挑選陽性轉化子,提取質粒并進行酶切驗證.如圖 1所示,經過 EcoRⅠ和 NotⅠ雙切,重組質粒產生約1600bp和 9300bp的兩條帶,與預計大小相符,證明重組質粒構建成功.
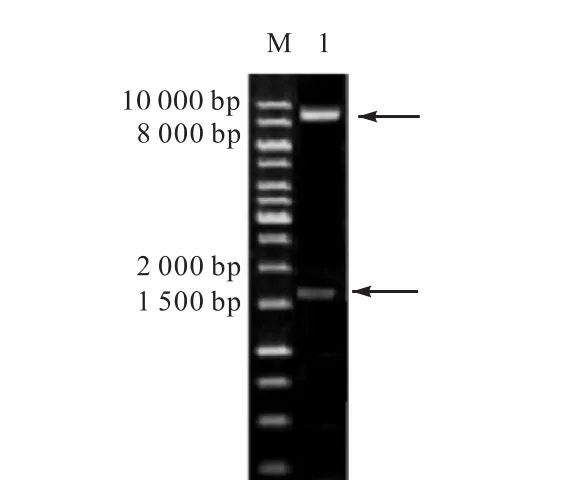
圖1 重組質粒的酶切鑒定Fig. 1 Enzyme digestion idenfication of recombinant plasmid
2.2 重組質粒電轉及轉化子鑒定
將重組質粒 pPIC9K-lacm線性化后電轉畢赤酵母GS115感受態,將電轉液涂布于MD平板.從MD平板上挑選轉化子,提取基因組,進行 PCR驗證.如圖2所示,得到一條約1600bp的條帶,證明lacm插入到畢赤酵母染色體中,成功構建重組菌株GS115/pPIC9K-lacm.
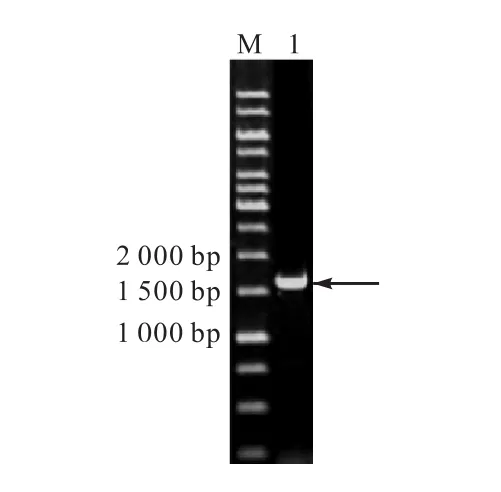
圖2 GS115/pPIC9K-lacm的PCR擴增結果Fig. 2 PCR amplification results of GS115/pPIC9K-lacm
2.3 rLACM的誘導表達及純化
對照菌株 GS115/pPIC9K和重組菌 GS115/pPIC9K-lacm經甲醇誘導發酵 120h后,離心得到發酵上清液.以2,6-DMP為漆酶底物時,測得rLACM的酶活力達到 0.37U/mL,而嗜熱棲熱菌來源的熱穩定性好的漆酶酶活力約為 0.022U/mL,嗜鹽菌Chromohalobacter salexigens來源的漆酶活力達到0.14U/mL[25,29].
對 rLACM 進行 SDS-PAGE分析,如圖 3(a)所示,GS115/pPIC9K-lacm 與 GS115/pPIC9K 相比,在相對分子質量 5.8×104左右多出一條帶,與預期rLACM 大小一致,以上說明 GS115/pPIC9K-lacm成功高效分泌表達rLACM.
如圖 3(b)所示,使用鎳柱親和層析法純化rLACM,并進行 SDS-PAGE分析,有一條單一條帶,其相對分子質量約為 5.8×104,可用于后續的酶學性質研究.
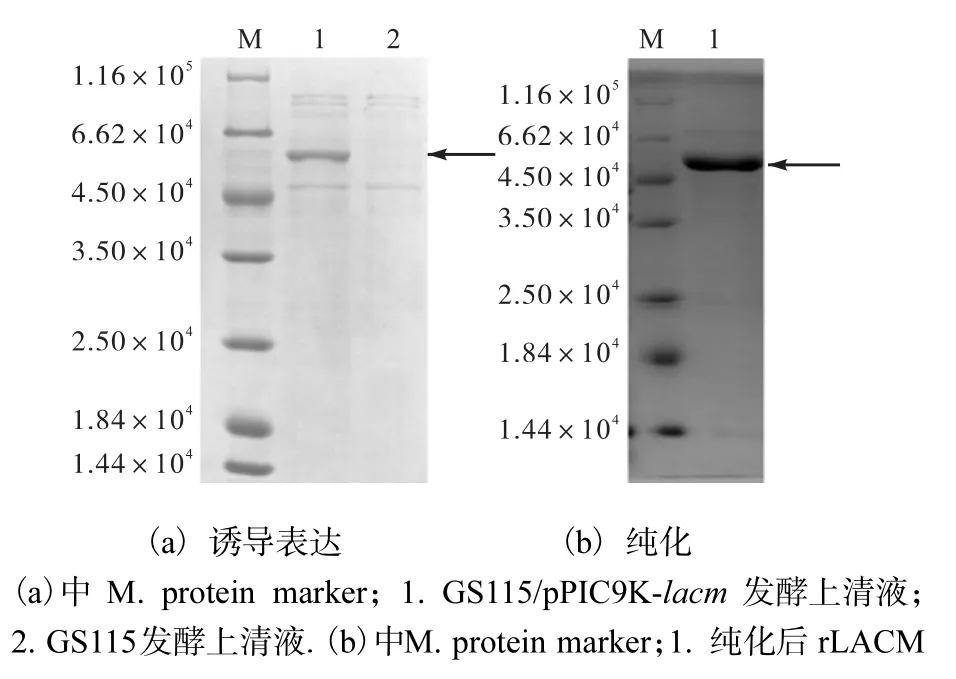
圖3 rLACM的SDS-PAGE分析Fig. 3 SDS-PAGE analysis of the rLACM
2.4 rLACM的酶學性質
2.4.1 rLACM的最適溫度和溫度穩定性
rLACM 的最適溫度和溫度穩定性如圖 4所示.rLACM 的最適反應溫度為 70℃,在 30~90℃范圍內具有酶活性.對 rLACM 的熱穩定性進行分析,其在30℃和40℃條件下保溫5h,酶活力可維持在 95%左右;在 50℃和 60℃條件下保溫 5h,酶活力仍在 50%以上;在 80℃條件下保溫 1h,殘余活力為25%左右.

圖4 rLACM的最適溫度和溫度穩定性Fig. 4 Optimum temperature and thermostability for rLACM
由此可見,rLACM 的熱穩定性能比真菌中的米曲霉(Aspergillus oryzae)以及細菌中的嗜水氣單胞菌(Aeromonas hydrophila)的漆酶熱穩定性能更加穩定[30-31].另外,黃俊等[20]報道的肺炎克雷伯氏菌 601(Klebsiella sp.601)來源漆酶在 80℃處理 30min,則完全失活;而 rLACM 在 80℃條件下保溫 1h,殘余活力為 25%左右.這表明 rLACM 具有較好的熱穩定性.
2.4.2 rLACM的最適pH和pH穩定性
rLACM的最適 pH和 pH穩定性如圖 5所示.rLACM的最適pH為8.0,在pH 7.0~9.0的范圍內具有酶活性.對 rLACM 的 pH穩定性進行分析,其在pH 7.0和pH 8.0的條件下保溫35h,其殘余活力在70%以上.在pH 5.0和pH 6.0以及pH 8.0和pH 9.0的條件下保溫20h,其殘余活力在60%以上,而黃俊等[20]報道 Klebsiella sp.601來源漆酶在 pH 8.0和 pH 9.0條件下保溫 24h殘余酶活力分別為55%和10%左右.此外,在pH 4.0、pH 10.0的條件下保溫20h,其殘余活力仍分別保持在41%和35%.該結果表明,rLACM具有較寬的pH穩定范圍.
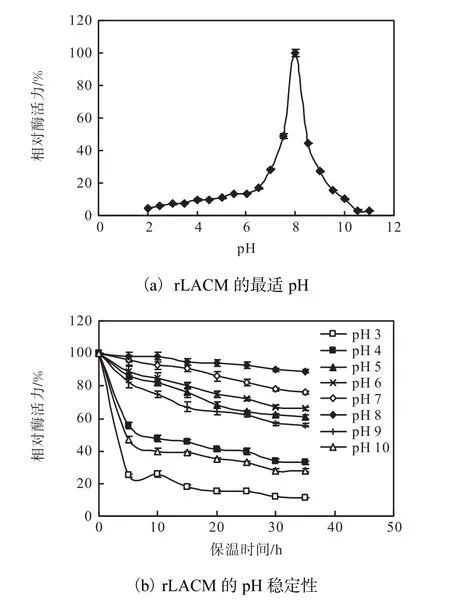
圖5 rLACM的最適pH和pH穩定性Fig. 5 Optimum pH and pH stability for rLACM
由上述研究表明,rLACM 與目前報道的細菌來源漆酶相比,兼具較好的熱穩定性及較寬的 pH穩定范圍;同時,經在畢赤酵母中表達 lacm基因獲得的rLACM,其酶學性質與在大腸桿菌中表達的重組漆酶[24]相似,說明表達宿主的改變并未影響漆酶的特性,從而可以通過畢赤酵母表達系統制備該性能優良的rLACM.
3 結 語
本研究以肺炎克雷伯氏菌的基因組為模板,通過PCR擴增得到其漆酶基因 lac,基于畢赤酵母密碼子偏愛性優化后得到新型漆酶基因 lacm,并成功構建重組菌株 GS115/pPIC9K-lacm,以 2,6-DMP為底物,發酵液中的酶活力達到 0.37U/mL,實現肺炎克雷伯氏菌漆酶的高效分泌表達.經酶學性質分析表明,以 2,6-DMP為底物時,rLACM 的最適溫度為70℃,最適pH為8;在30~70℃、pH 5.0~9.0活力穩定.該rLACM表現出的良好的酶學特性為漆酶的應用奠定了基礎.
