冬小麥冠層-大氣氨交換的季節性特征及其影響因素
2019-02-25 10:13:00景建元李惠通鄭天義呂慎強周春菊王林權
干旱地區農業研究 2019年6期
關鍵詞:生長
王 呂,景建元,李惠通,鄭天義,李 嘉,呂慎強,周春菊,王林權
(1.西北農林科技大學資源環境學院,陜西 楊凌 712100;2. 中國農業科學院農業資源與農業區劃研究所,北京 100081;3. 西北農林科技大學生命科學學院,陜西 楊凌 712100)
農業生產活動是大氣氨(NH3)的主要排放源,其中施肥引起的NH3排放可達40%[1]。大氣中的NH3可與空氣中的NOx、SO2等發生反應生成PM2.5的主要成分[2],誘發灰霾;日益增加的大氣氮沉降,威脅著陸地和水體生態系統[3-5]。農田NH3排放已經從一個農業肥料效應問題轉化成了一個生態環境問題。關于土壤和作物生育后期冠層NH3揮發對農田NH3排放的貢獻已有大量的研究報道[6-8],但作物整個生育期冠層-大氣氨交換對農田NH3揮發的貢獻還缺乏系統研究。有報道指出[9]:高氮處理(180 kg·hm-2)的冬小麥灌漿前存在冠層吸收NH3的現象,灌漿后存在NH3的釋放,兩者占同時期土壤NH3揮發的0.1%~9.7%和65.1%~74.5%。從整個生育期看,冬小麥和水稻冠層具有消減農田氨揮發的作用[10]。也有學者認為作物冠層可以吸收外界大氣[11]和土壤[12]中揮發的NH3,但作物冠層吸收環境中的NH3在提高氮肥利用率的同時,也可能產生氨毒害作用。因此要正視作物冠層NH3交換規律、冠層NH3交換對農田氨揮發貢獻的時空變化規律以及季節性氨排放系數的效應。

1 材料與方法
1.1 試驗設計
試驗在西北農林科技大學農作一站(2016—2017)和曹新莊試驗農場(2017—2018)進行,試驗區域屬半濕潤易旱區,耕層土壤理化性質如下表1。供試材料選用小偃22(XY22)和鄭麥366(ZM366),播種量為120 kg·hm-2。供試肥料為尿素(含N 46%),試驗設置不施氮和施氮180 kg·hm-2兩個氮肥水平。試驗共4個處理,分別為小偃22不施氮(XY22-N0),小偃22施氮180 kg·hm-2(XY22-N180),鄭麥366不施氮(ZM366-N0),鄭麥366施氮180 kg·hm-2(ZM366-N180),每個處理重復3次。返青期加設去除枯葉黃葉處理(代號N0-Q和N180-Q)。小區面積為6 m×2.6 m=15.6 m2。播種前將2/3尿素和磷肥(重過磷酸鈣:含P2O546%,90 kg P2O5·hm-2)均勻撒施到地表,翻耕混勻,平整地面。各小區按25 cm行距開溝播種。播種當天即安放NH3揮發收集裝置。將剩余1/3的尿素于拔節期雨前溝施追肥。
1.2 采樣與測定
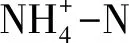

表1 耕層土壤(0~20 cm)理化性質
去除枯葉黃葉處理:即返青之后把冬小麥葉片有黃葉的部分和干枯的葉片剪去。成熟之后刈割小麥留20 cm麥茬,對麥茬繼續測定NH3揮發,持續15 d。
冬小麥各生育期時間段分別對應為:分蘗期(10月初~12月下旬),越冬期(12月下旬~翌年2月底),返青期(3月上旬)、拔節期(3月中、下旬)、孕穗~揚花期(4月)、灌漿期(5月)、成熟期(6期上、中旬)。
參照文獻[9]不同高度下層海綿的NH3回收率對吸收的NH3進行換算。參照文獻[9]計算土壤NH3揮發速率(kg·hm-2·d-1)、冠層NH3揮發速率(kg·hm-2·d-1)、土壤NH3揮發累積量、冠層NH3揮發累積量、麥田NH3揮發累積量。
枯葉黃葉NH3揮發累積量(kg·hm-2)
ALAV=∑(MN-M(N-Q))/(S×R)×10 000
麥茬NH3揮發累積量(kg·hm-2)
AWSAV=∑(M(N-WS)-M(CK- N))/(S×R)×10 000
其中,MN為不去除枯葉黃葉冠層NH3揮發量,M(N-Q)為去除枯葉黃葉冠層NH3揮發量,M(N-WS)為遮罩麥茬NH3揮發量,M(CK- N)為不遮罩植物NH3揮發量,S為收集裝置的橫截面積,R為不同海綿高度氨捕獲裝置的氨氣回收率。ALAV為正值說明枯葉黃葉參與了冠層氨揮發,負值表示枯葉黃葉沒有參與冠層氨揮發過程。

1.3 數據處理
數據應用Excel 2010程序和SAS 8.1統計分析軟件進行處理,使用雙因素完全隨機區組設計方差分析和最小顯著性差異法( LSD) 對數據均值進行多重比較,回歸分析前將有關數據參照文獻[20]做標準化處理,Origin2015進行繪圖。
2 結果與分析
2.1 冠層NH3揮發動態
2016—2017年和2017—2018年冬小麥生長季,基肥和追肥后15 d內均出現土壤NH3揮發速率峰值,與峰值對應的冠層NH3揮發速率為負值,說明有冠層NH3吸收現象,追肥后(拔節期)土壤NH3揮發和冠層NH3吸收比基肥(分蘗期)更為明顯。2016—2017年生長季越冬期,兩品種的土壤NH3揮發速率小,無明顯冠層NH3吸收現象;2017—2018年生長季越冬期土壤NH3揮發速率明顯,冠層NH3揮發速率為負值,冠層有NH3吸收。返青期各處理土壤NH3揮發速率也有一個峰值,相應的冠層NH3揮發速率為負值,說明返青期也有冠層氨吸收,小偃22冠層NH3吸收較鄭麥366明顯。孕穗~揚花期,施肥處理冠層NH3揮發速率為負值,有NH3吸收。到灌漿~成熟期,土壤NH3揮發下降并維持相對穩定,冠層NH3揮發速率以正值為主,表現出明顯的冠層NH3釋放,且2017—2018年生長季的土壤NH3揮發速率與冠層氨交換均顯著高于2016—2017年生長季。
2.2 冠層氨釋放對麥田氨揮發的貢獻及其品種與施肥效應
2016—2017年生長季,品種和氮肥均對冠層和麥田NH3揮發累積量有顯著影響(P<0.1)。小偃22全生育期以NH3吸收為主,鄭麥366為NH3釋放,兩品種差異顯著(P<0.1)(表2)。 氮肥處理麥田NH3揮發累積量較不施氮高1.840 kg·hm-2,達差異顯著水平(P<0.1)。
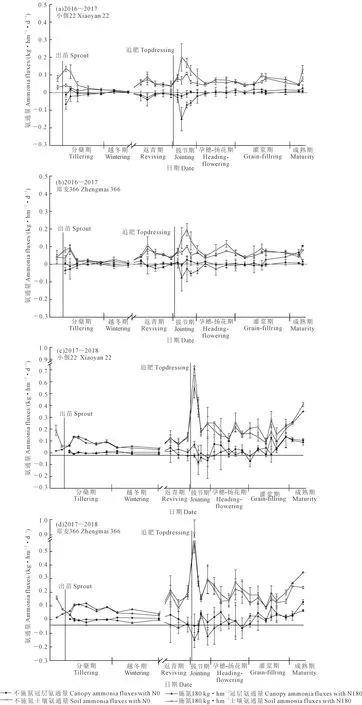
圖1 2016—2017年和2017—2018年生長季NH3通量變化特征Fig.1 Dynamics of ammonia fluxes of growing seasons of 2016-2017 and 2017-2018
由表3可得,2017—2018年生長季,品種顯著影響冠層NH3揮發累積量(P<0.1),小偃22冠層為NH3釋放,鄭麥366為吸收。施氮對麥田氨揮發也有顯著影響(P<0.1)。
2016—2017年生長季,在分蘗期和拔節期,施氮處理冠層NH3吸收累積量為0.216~1.130 kg·hm-2,可消減17%~120%麥田NH3揮發,鄭麥366冠層NH3吸收量較小偃22低70.1%和37%。返青期施氮處理的冠層可消減17%~32%麥田NH3揮發,且鄭麥366吸收量比小偃22低32%。孕穗~揚花期,除鄭麥366不施氮處理外,其它處理均表現為冠層NH3的吸收,其中鄭麥366施氮處理冠層NH3的吸收量為麥田NH3揮發量的22%。在灌漿期和成熟期,施氮處理冠層NH3揮發累積量為0.262~0.820 kg·hm-2,對麥田的氨揮發的貢獻為14%~41%;不施氮為0.015~0.096 kg·hm-2,貢獻為4%~9%。
2017—2018年生長季,分蘗和拔節期,施氮處理冠層NH3吸收累積量為0.144~1.306 kg·hm-2,消減1%~12%麥田NH3揮發。越冬期,小偃22不施氮處理冠層NH3吸收量達麥田NH3揮發量27%,鄭麥366施氮處理為23%。返青期和孕穗~揚花期,鄭麥366施氮處理冠層NH3吸收占麥田NH3揮發的比例為36%。在灌漿和成熟期,各處理均表現為冠層NH3的釋放,施氮處理冠層NH3揮發累積量為0.317~1.204 kg·hm-2,不施氮處理為0.526~1.615 kg·hm-2;施氮處理和不施氮處理的冠層NH3揮發對麥田氨揮發的貢獻分別為7%~24%和16%~28%。
綜合兩個冬小麥生長季,小偃22和鄭麥366分蘗期和拔節期冠層NH3吸收量對麥田NH3揮發的削減作用可達23%~40%。返青期,施氮處理的冠層NH3吸收量對麥田NH3揮發的削減作用可達17%~40%。灌漿~成熟期,各處理均為冠層NH3的釋放,施氮處理冠層NH3釋放對麥田NH3揮發的貢獻為17%~27%。
2.3 枯葉黃葉NH3揮發累積量
兩個冬小麥生長季在拔節期枯葉黃葉NH3揮發累積量均表現一致(見圖2),小偃22施肥處理與不施肥處理枯葉黃葉NH3揮發累積量均為負值,說明枯葉黃葉不是造成小偃22冠層NH3揮發的原因;而鄭麥366均為正值,說明鄭麥366枯葉黃葉參與了冠層NH3揮發。孕穗~揚花期,2016—2017年生長季各處理枯葉黃葉NH3揮發累積量均為正值,但累積量很少;而2017—2018年生長季,各處理NH3揮發累積量均為負值,說明枯葉黃葉未參與冠層NH3揮發。灌漿期,兩個冬小麥生長季表現出相反的特征,2016—2017年生長季,各處理枯葉黃葉NH3揮發累積量均為負值,說明枯葉黃葉未參與冠層NH3揮發;而2017—2018年生長季,各處理枯葉黃葉NH3揮發累積量均為正值,說明枯葉黃葉參與了冠層NH3揮發。

表2 品種和氮肥對2016—2017年生長季冠層及麥田NH3揮發累積量的影響/(kg·hm-2)

表3 品種和氮肥對2017—2018年生長季冠層及麥田NH3揮發累積量的影響/(kg·hm-2)
2.4 收獲后麥茬NH3揮發累積量
2016—2017年生長季,除小偃22不施氮處理,其它處理的麥茬均未發現NH3揮發;施氮處理土壤NH3揮發累積量均高于不施氮處理且差異顯著(P<0.05)。2017—2018年生長季,各種處理的麥茬NH3揮發量在0.093~0.535 kg·hm-2。施氮處理麥茬NH3揮發累積量和土壤NH3揮發累積量均顯著高于不施氮處理(P<0.05)。2017—2018年生長季土壤NH3揮發累積量均高于2016—2017年生長季(表4)。
2.5 影響冠層NH3通量的因素
2.5.1 冠層NH3揮發與耕層土壤溫度 冬前,隨著耕層土壤溫度逐漸降低,小偃22和鄭麥366冠層NH3揮發速率均有下降趨勢(如圖3a);冬后隨耕層土壤溫度升高,冠層NH3揮發速率升高,二者呈極顯著正線性關系(P<0.01)(如圖3b)。
2.5.3 NH3揮發速率、葉片硝酸鹽含量和硝酸還原酶活性 小偃22和鄭麥366冠層NH3揮發速率與硝酸鹽含量呈負線性關系,且小偃22關系達顯著水平(P<0.05)(圖5a)。小偃22和鄭麥366冠層NH3揮發速率與硝酸還原酶活性無顯著相關性(圖5b)。

注:N0代表不施氮遮罩植株處理NH3揮發與遮罩去除枯葉黃葉植株處理NH3揮發的差值之和,N180代表施氮180 kg·hm-2遮罩植株處理NH3揮發與遮罩去除枯葉黃葉植株處理NH3揮發的差值之和。同一生育時期不同小寫字母表示同類因素的處理間差異顯者(P<0.05)。Note: N0, the sum of the difference between NH3 volatilization in the treatment of NH3 volatilization and the removal of NH3 volatilization from the dry and yellow leaf plants without nitrogen treatment; N180, the sum of the difference between NH3 volatilization in the treatment of NH3 volatilization and the removal of NH3 volatilization from the dry and yellow leaf plants with nitrogen application of 180 kg·hm-2. Values followed by different lowercase letters during same growth period represent significantly difference between the treatments at the 0.05 level for the same factor.圖2 枯葉落葉NH3揮發累積量Fig.2 Ammonia volatilization of dry and yellow leaves
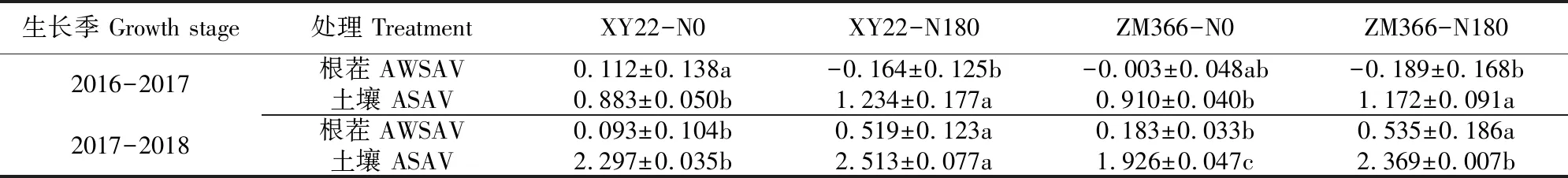
表4 收獲后15 d麥茬和土壤NH3揮發累積量/(kg·hm-2)

注:*,**分別表示在0.05,0.01水平上差異顯著。Note: *,** indicate significantly difference at 0.05 and 0.01 level, respectively, the same below.圖3 冠層NH3通量與耕層土壤溫度的關系Fig.3 Relationship between canopy ammonia fluxes and tillage soil temperature
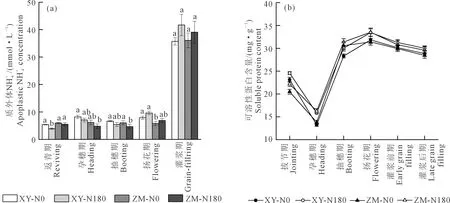
注:同一生育期不同小寫字母表示同類因素的處理間差異顯著(P<0.05)。Note: Values followed by different lowercase letters during the same growth period indicate significantly difference between the treatments at the 0.05 level for the same factor.圖4 葉片質外體濃度、可溶性蛋白含量Fig.4 Apoplastic concentration and soluble protein content of leaves
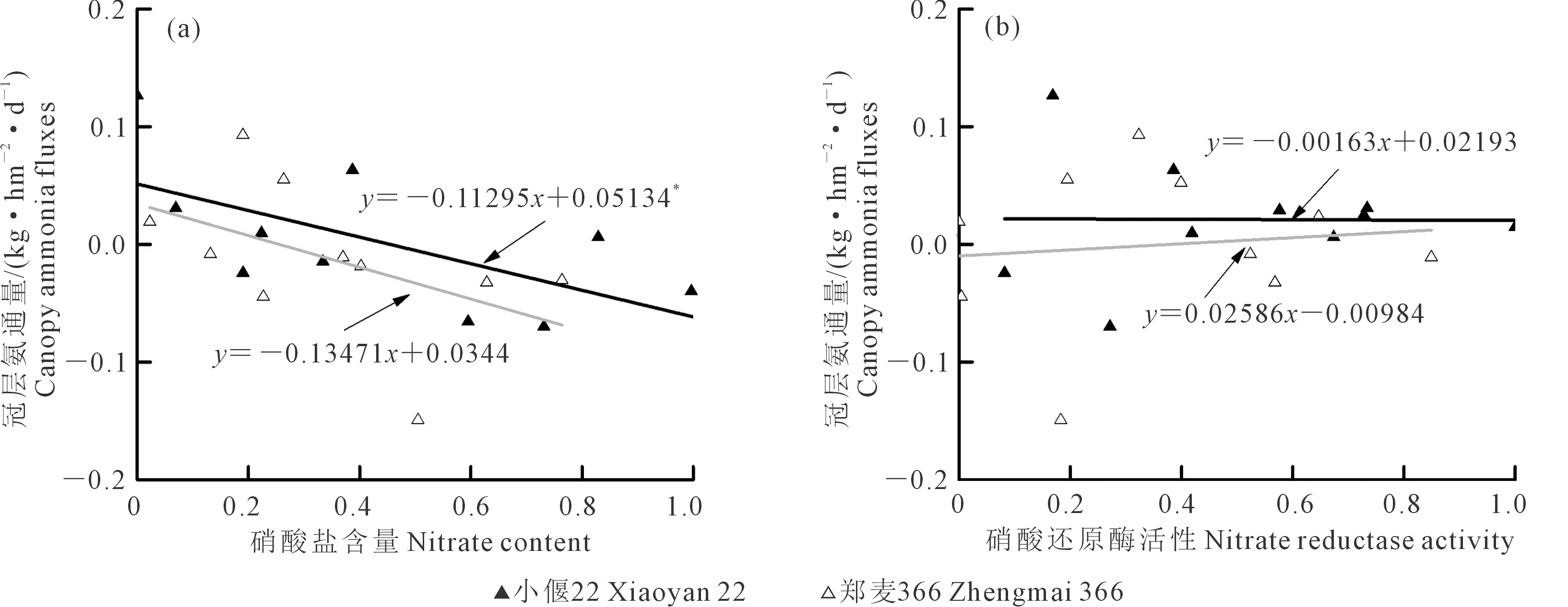
圖5 冠層NH3通量與硝酸鹽含量和硝酸還原酶活性的關系Fig.5 Relationship between canopy ammonia fluxes and nitrate content with nitrate reductase activity
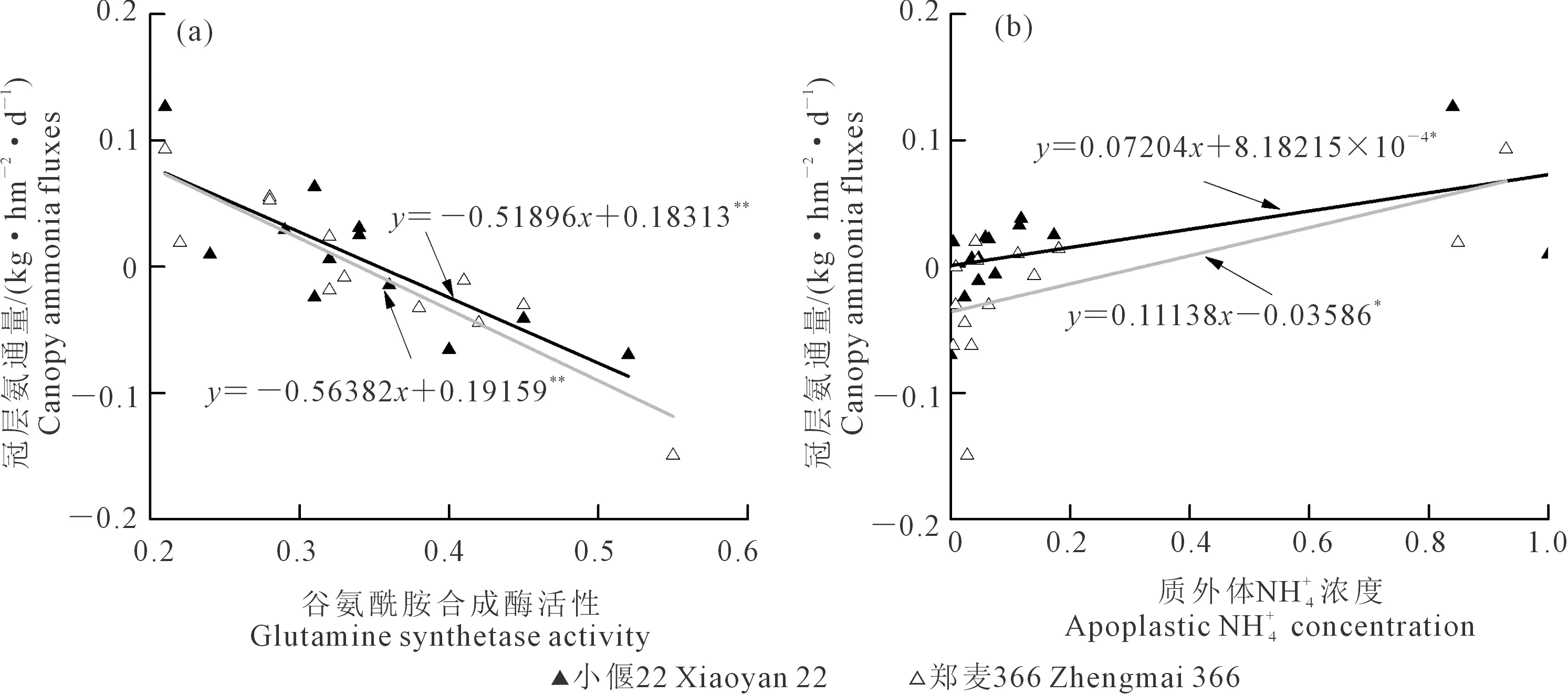
圖6 冠層NH3通量與谷氨酰胺合成酶活性和質外體濃度的關系Fig.6 Relationship between canopy ammonia fluxes and glutamine synthetase activity with apoplastic concentration
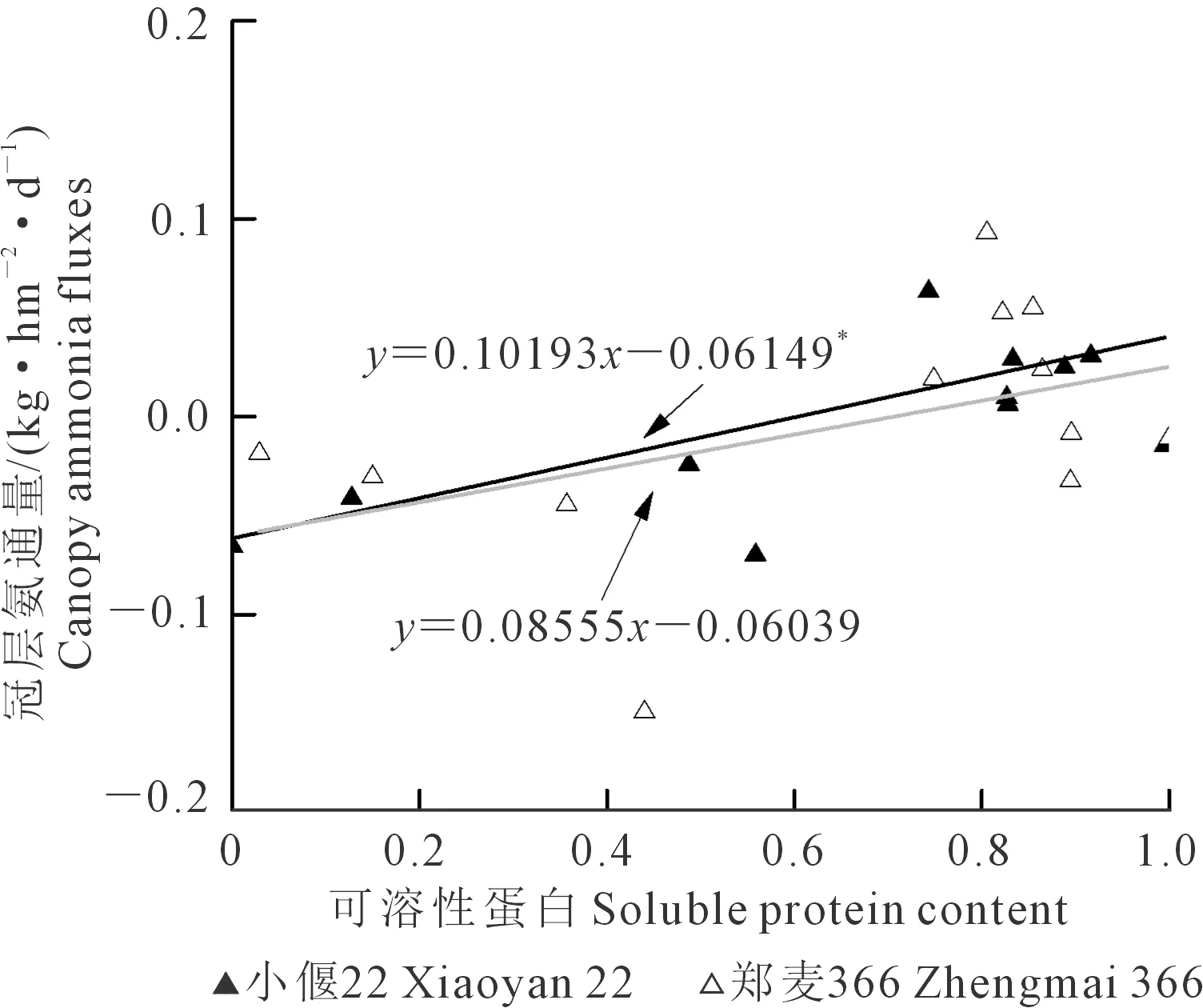
圖7 冠層NH3通量與可溶性蛋白含量的關系Fig.7 Relationship between canopy ammonia fluxes and soluble protein content
3 討 論
3.1 冠層NH3交換的季節性分布
冬小麥冠層NH3交換在不同生育期表現出不同的規律,分蘗期(10—11月)、返青(2—3月初)和拔節期(3月中下旬)冠層是氨匯,冠層NH3吸收對減少麥田NH3揮發的貢獻為23%~40%。這與前人研究[21]關于冠層可直接吸收大氣中的NH3,與農田接壤的森林也會吸收農田釋放的NH3[22]結論一致。在越冬期(12月~翌年2月上中旬),土壤和冠層NH3揮發較少。因此,營養生長期小麥會主動吸收空氣中的NH3[23]。孕穗~揚花期(4月)施氮處理均為冠層NH3的吸收,但吸收量較少。到灌漿~成熟期(5月),土壤NH3揮發下降且維持在一定低水平,冠層為NH3揮發。這與生育后期冠層NH3揮發特性的報道一致[9,24]。籽粒收獲后2周(6月初),麥茬也存在NH3釋放,籽粒收獲后15 d內麥茬可累計釋放的NH3為0.093~0.535 kg·hm-2。特別是在高肥力和施用氮肥條件下較顯著。因此收獲后土壤和根茬的氨揮發不容忽視。
3.2 枯葉黃葉NH3揮發
枯、黃葉對冠層NH3揮發的影響受冬小麥品種、生育時期和土壤肥力等的影響。在拔節期鄭麥366枯枝落葉參與冠層NH3揮發,而小偃22則無,這可能與小麥品種間的生長發育進程及氮代謝的差異有關,如小偃22植株比鄭麥366高10 cm,比鄭麥366早熟3~7 d,葉片為細長型,而鄭麥366為寬厚型。在孕穗~揚花期,小偃22和鄭麥366枯葉落葉均未參與冠層NH3揮發,可能是此生育期營養生長和生殖生長對氮代謝需求旺盛,老葉氮素向穗器官轉運速度較快。灌漿期,小偃22和鄭麥366在2017—2018年生長季,枯葉黃葉參與了冠層NH3揮發,而2016—2017年生長季沒有,原因可能與土壤肥力和植株氮素營養水平有關。2017—2018年生長季,播前土壤礦質氮含量幾乎為2016—2017年生長季的2倍,根系吸收的氮素對籽粒的貢獻較大,而葉片氮素向籽粒的轉運下降,使得這部分養分來不及轉運至籽粒而造成冠層NH3揮發的增加。2016—2017年灌漿期小偃22和鄭麥366施氮處理葉片氮素累積量分別為2.36 mg·莖-1和0.90 mg·莖-1,不施氮處理分別為0.49 mg·莖-1和0.34 mg·莖-1,2017—2018年生長季小偃22和鄭麥366施氮處理葉片氮素累積量為5.34 mg·莖-1和2.97 mg·莖-1,不施氮處理為4.15 mg·莖-1和2.50 mg·莖-1(未發表)。有研究指出冬小麥器官脫落物中的氮含量僅占作物生長后期減少總量的4%左右[25],還有學者利用同位素15N觀測植物根系和根系周圍的土壤并沒有發現冠層氮素遷移分泌的蹤跡[26]。因此,枯黃葉對冠層氨揮發的貢獻有限。
3.3 冠層NH3揮發的農學因素
NH3揮發與土壤溫度密切相關[27]。冬前(10—12月)隨著氣溫的降低,冠層活動減弱,因此溫度與冠層氨揮發的關系不密切。冬后(3—6月)隨著溫度升高,土壤氮素轉化和作物冠層活動趨于旺盛,耕層土壤溫度與冠層NH3揮發極顯著正相關(圖3),冠層NH3的吸收和釋放隨施氮量的增加而增加,施氮處理冠層NH3吸收是不施氮處理的5.3~10.3倍,冠層NH3釋放為0~8.5倍(2016—2017年生長季)。冠層NH3的吸收和釋放也受土壤肥力的影響,小麥一生中1/3氮素吸收來源于肥料氮,2/3來源于土壤氮[28]。高肥力有利于土壤氨揮發及其與冠層氨交換。2017—2018年生長季,播前土壤礦質氮含量是2016—2017年生長季的2倍,不施氮處理冠層NH3揮發是施氮處理的1.2~2.1倍。2016—2017年生長季,麥茬NH3揮發不明顯(除小偃22施氮處理),而2017—2018年生長季,麥茬NH3揮發明顯,這也與土壤肥力和氮素供應導致麥茬含氮量高有關。2016—2017年生長季小偃22和鄭麥366施氮麥茬氮素含量分別為2.88 g·kg-1和2.05 g·kg-1,不施氮處理為1.08 g·kg-1和1.20 g·kg-1,2017—2018年生長季小偃22和鄭麥366施氮處理麥茬氮素含量分別為7.62 g·kg-1和5.25 g·kg-1,不施肥處理分別為6.09 g·kg-1和5.14 g·kg-1(未發表)。2017—2018年生長季,充足的土壤氮素供應使得收獲后麥茬含有比較多的氮素,為氨揮發提供了氮源。
3.4 影響冠層NH3揮發的生理因素

4 結 論
1)冬小麥冠層NH3的吸收和釋放因生育期而異,孕穗~揚花期之前以冠層NH3吸收為主,對麥田的消減率為23%~40%;孕穗~揚花期之后以冠層NH3釋放為主,對麥田的貢獻率為17%~27%。施氮處理和高土壤肥力條件下的冠層NH3釋放和吸收均較不施肥或低土壤肥力大。
2)冬小麥枯葉黃葉對冠層NH3揮發的影響較小,且受生育期、小麥品種及土壤肥力等影響。
3)冠層氨揮發受環境條件影響,與植株氮代謝密切相關。冠層氨釋放與冬后土壤溫度、質外體銨濃度、可溶性蛋白顯著正相關,而谷氨酰胺合成酶活性和葉片硝酸鹽含量顯著負相關。營養生長期的冠層氨交換受氨同化過程的控制,灌漿末期的冠層氨釋放受葉片等器官的氮素轉運過程控制。
猜你喜歡
祝您健康·文摘版(2024年6期)2024-07-26 00:00:00
小讀者(2021年2期)2021-03-29 05:03:48
少兒美術(2020年3期)2020-12-06 07:32:54
現代裝飾(2020年11期)2020-11-27 01:47:48
中學生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年級(2020年3期)2020-03-17 09:24:33
瘋狂英語·新悅讀(2019年11期)2019-12-18 05:14:16
華人時刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
