番茄SlETR6基因的克隆及非生物脅迫下的表達分析
2019-03-08 02:00:10蘇麗艷
華北農學報 2019年1期
關鍵詞:植物
蘇麗艷
(西安文理學院 生物與環境工程學院,秦嶺野生觀賞植物研究中心, 陜西 西安 710065)
乙烯是一種小分子氣態植物激素,對果實成熟、種子萌發、葉和花衰老等多種植物發育過程有重要調控作用[1-2]。此外,乙烯在植物響應生物和非生物脅迫過程中有重要的作用,當植物體受到冷害、干旱、損傷、低氧等生物及非生物逆境脅迫時,其體內乙烯的含量往往會增加[3]。乙烯信號通過相應的轉導途徑進行脅迫信號的傳遞,調控下游基因的表達進而使植物體在生理水平上發生一系列適應環境的改變。乙烯受體 (Ethylene receptor, ETR) 蛋白是乙烯的負調控因子,是乙烯信號轉導途徑上游重要組成元件之一,在缺乏乙烯時,受體會抑制乙烯誘導的相關基因表達;乙烯存在時,乙烯與受體蛋白結合使受體失活,從而激活下游的乙烯誘導相關基因[4-5]。
ETR家族包含多個同源基因,擬南芥中分離得到5個ETR受體基因,根據其結構域可分為兩大亞家族,ETR1亞家族包括AtETR1和AtERS1,ETR2亞家族包括AtETR2、AtERS2、AtEIN3[6]。ETR1亞家族含有組蛋白激酶所需要的5個完整結構域,而ETR2亞家族基因僅含有部分組蛋白激酶所需要的結構域。番茄中分析篩選到6個乙烯受體,分別是:SlETR1、SlETR2、NR(Never-ripe)、SlETR4、SlETR5和SlETR6,它們在蛋白質水平上差異較大,在序列上表現出50%以下的同源性[7-10]。目前已經在鳳梨、蘋果、草莓、桃及芒果等多種植物中被分離出來[11-16]。
目前,對于乙烯受體ETR的研究多集中在其對植物生長發育的調控作用,如番茄乙烯受體SlETR1和SlETR2在乙烯信號轉導中起負調控作用[17-18];NR突變體果實不能正常成熟[19];SlETR4的存在可促使番茄果實乙烯響應系統Ⅱ中乙烯的加速合成,促進果實早熟[20-21],果實成熟期時,SlETR5的表達明顯上升,而SlETR6的功能可能與SlETR4相似。目前,對于ETR在脅迫過程中的功能只有少量報道,前期研究發現,AtEIN2突變后影響擬南芥植株的鹽敏感度,證明AtEIN2參與鹽脅迫信號的調控過程[22];可可CcEIN4抑制表達后可以提高植物對鹽脅迫的抗性,表明乙烯受體CcEIN4亦參與調控植株的鹽脅迫信號調控過程[23]。
番茄是研究呼吸躍變型果實發育特征的典型模式植物。SlETR6是番茄乙烯受體家族中發現較晚的一個成員,目前尚無SlETR6對干旱、高鹽、溫度脅迫響應的相關報道。本研究對番茄矮化品種Mic-Tom中SlETR6基因進行了克隆,并研究了SlETR6在不同植物組織及高鹽、高溫、低溫、干旱等脅迫條件下的表達模式,旨在為進一步了解SlETR6在植物響應環境脅迫中的功能奠定理論基礎。
1 材料和方法
1.1 試驗材料及脅迫處理
Mic-Tom番茄種子為中科院遺傳所饋贈。種植于西安文理學院植物生長繁育單元-西北文絡型連棟溫室中,番茄生長條件:白晝溫度分別為光周期為16 h/8 h,晝夜溫度為27 ℃/19 ℃, 濕度為60%~80% RH。取番茄植株的不同植物組織,包括根、莖、葉、花、種子和不同發育時期果實(小果/花后5 d、中果/花后10 d、大果/花后25 d、轉色果/35 d、紅果/花后45 d),取樣后立即用液氮凍存并存于-80 ℃冰箱備用。
選取盆栽長勢一致的Mic-Tom番茄35 d苗植株進行非生物脅迫處理,包括200 mmol/L NaCl溶液灌根、高溫(40 ℃)、低溫(4 ℃)、脫水即模擬干旱(將番茄苗洗凈根部泥土置于濾紙上),以未做任何處理的植株為對照。每個處理3次重復,分別于處理后0,1,3,6,12,24 h收取葉片組織,液氮凍存后存于-80 ℃冰箱備用。
1.2 番茄總RNA提取及SlETR6基因的克隆
試驗材料總RNA提取使用天根生化科技有限公司RNA提取試劑盒, 用1%瓊脂糖凝膠電泳測定其濃度和完整性,利用DNase去除基因組DNA后,利用TaKaRa反轉錄試劑盒按照產品操作按說明書合成cDNA。
以番茄葉片cDNA為模板進行SlETR6基因ORF全長克隆,利用Primer 5.0設計引物SlETR6-F:5′-GCAATGATGAAGAAAGTA-3′,SlETR6-R:5′-GTC ATGGCATTCCTCTG-3′。采用TaKaRa 公司的PrimeSTAR?HS DNA Polymerase高保真酶進行擴增,反應體系為25 μL,反應程序是:94 ℃,5 min;94 ℃,30 s,58 ℃,30 s,72 ℃,90 s,34個循環; 72 ℃,10 min。PCR產物在1%瓊脂糖凝膠電泳檢測,將檢測到的目的片段用瓊脂糖凝膠純化回收,將回收到的目的基因產物連接到pMD19-T載體后,轉化大腸桿菌,篩選陽性克隆提取質粒后進行DNA測序檢測。
1.3 番茄SlETR6基因的生物信息學分析
利用 DNAMAN 對SlETR6基因進行氨基酸序列比對;蛋白質分子質量和等電點預測利用在線軟件 ProtParam(http://web. expasy.org/protparam/)計算完成;利用TargetP 1.1 Server、plantCARE 在線工具完成SLETR6蛋白的亞細胞定位預測、啟動子順式作用元件分析。利用 MEGA 5 軟件構建SlETR6基因系統進化樹。
1.4 SlETR6表達特性分析
對番茄不同植物組織及不同脅迫處理得到的材料提取RNA, 純化后反轉錄為cDNA。在ABI Quant Studiotm6 Flex Real-time PCR 系統進行qPCR 分析。以cDNA為模板,以SlUBI(GenBank登錄號:Q96483)為內參。用 Primer 5.0 設計引物,qSlETR6-F:5′-GAATGGCATCCACGGAG-3′,qSlETR6-R: 5′-CAATCTCCTCAGTGTCG-3′;SlUBI-F: 5′-TGT CCCTATCTACGAGGGTTAT-3′;SlUBI-R:5′-AGTTAA ATCACGACCAGCAAGAT-3′。主要步驟按熒光定量試劑盒說明書操作完成。反應總體系為20 μL,10 μL 熒光染料反應混合液,1 μL 模板cDNA (濃度 30 ng/μL),引物各1 μL (10 μmol/L),ddH2O 7 μL。反應程序為:95 ℃變性3 min;然后,95 ℃ 20 s,60 ℃退火30 s,72 ℃延伸30 s,共39次循環。qRT-PCR 反應于ABI Quant Studiotm6 Flex Real-Time PCR System上進行。相對表達量用2-ΔΔCT法分析[24]。
2 結果與分析
2.1 SlETR6基因的克隆和啟動子序列分析
利用PCR方法克隆得到2 200 bp左右與預期大小一致的目標片段(圖1)。測序結果表明,目標條帶大小2 265 bp,與引物設計預測條帶一致,編碼754個氨基酸,其組成蛋白質分子質量為85.05 ku, 等電點為7.28。啟動子序列分析表明,SlETR6含有TATA 框、CAAT 框等調控真核基因轉錄的基本元件外,還具有熱脅迫、干旱脅迫、低氧脅迫、光響應、激素(乙烯、水楊酸、赤霉素)誘導響應、防御與脅迫響應元件等多種功能響應元件(表1)。
2.2 SlETR6同源序列對比及進化樹分析
利用BlastP對SlETR6基因編碼的氨基酸同源性進行檢索,番茄SlETR6基因編碼的氨基酸與花生AiETR2蛋白(XP_016175314.1)、向日葵HaETR2-like蛋白(XP_022002894.1)、胡桃JrETR2-like蛋白(XP_018810721.1)、苦瓜McETR2蛋白(XP_022146885.1)、木薯MeETR2-like蛋白(XP_021607133.1)、煙草NaETR2-like蛋白(XP_019227687.1)、櫻桃PaETR2-like蛋白(XP_021807404.1)、矮牽牛PhETR蛋白(AAZ81985.1)、梅PmETR2(XP_008224393.1)、桃PpETR2蛋白(XP_007221962.1)、蓖麻RcETR2蛋白(XP_002529316.1)、芝麻SiETR2(XP_011091175.1)、馬鈴薯StETR-like蛋白(XP_006354517.1,)、棗ZjETR2蛋白(XP_015887972.1)等具有較高的相似性(圖2)。系統進化樹分析表明,其中番茄SlETR6與馬鈴薯StETR-like蛋白的同源性最高(圖3)。

M. Marker Ⅲ DNA 分子標記;SlETR6. PCR 產物。M. Marker Ⅲ DNA Marker; SlETR6. PCR product.

表1 SlETR6啟動子序列順式元件分析Tab.1 The analysis of putative cis-elements in the promoter of SlETR6 genes


圖2 番茄SlETR6與其他相似性高的氨基酸聚類分析Fig.2 Phylogenetic tree of amino acid sequences of SlETR6 of tomato and other similar amino acids

圖3 番茄SlETR6 與其他植物氨基酸序列的進化樹分析Fig.3 Phylogenetic tree of SlETR6 and other plant species
2.3 SlETR6基因的組織表達分析
qPCR檢測發現,SlETR6在番茄根、葉、花、果實中均有表達,其在花中相對表達量較高,在葉中最低,在花、紅果和葉片中的表達量與種子相比存在顯著差異(圖4-A);對SlETR6在果實不同發育時期的檢測發現,轉色果中表達量最高,紅果和大果次之,在轉色果、紅果和大果中的表達量與小果相比存在顯著性差異(圖4-B)。
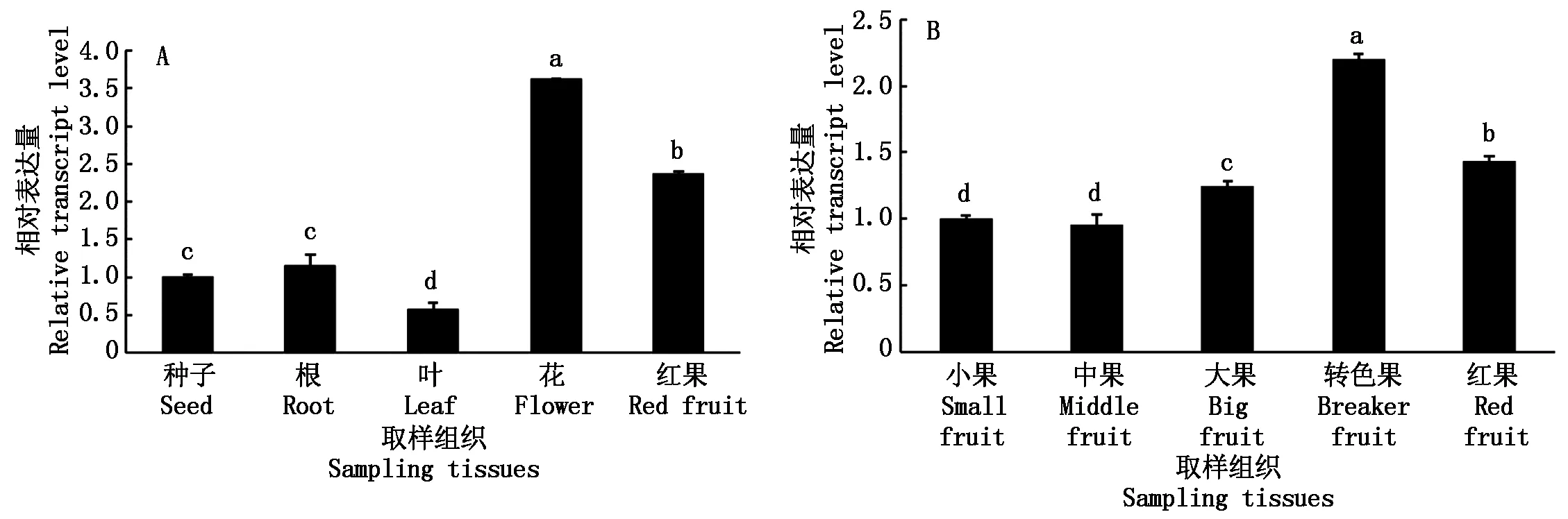
A.不同植物組織中SlETR6基因的表達情況;B.果實發育不同時期SlETR6基因的表達情況。不同小寫字母表示在0.05水平差異顯著。圖5-7同。A. Relative expression of SlETR6 in different tissues; B. Relative expression of SlETR6 at different stage of fruit development. Different small letters show significantly different at 0.05.The same as Fig.5-7.
2.4 SlETR6基因在高鹽脅迫下的表達分析
為了解SlETR6基因對高鹽響應的表達特征,利用qPCR檢測了其在脅迫后不同時間段的表達情況。結果顯示:高鹽脅迫處理6,12 h后SlETR6相對表達量具有顯著性提高,且在處理后12 h表達量達到最高,相對表達量約為對照組的3.6倍,損傷處理24 h后SlETR6基因的相對表達量恢復正常水平(圖5)。
2.5 SlETR6基因在溫度脅迫下的表達分析
如圖6所示,高溫 (40 ℃) 儲藏后SlETR6基因表達量表現出先升高(1~3 h)后下降(6~12 h)的趨勢,并在高溫處理3 h后達到峰值。高溫脅迫24 h后其相對表達量恢復正常水平。低溫4 ℃脅迫處理后,SlETR6基因相對表達量在低溫處理后1,6,12,24 h表現出下調趨勢。

圖5 高鹽脅迫下SlETR6基因的表達情況Fig.5 Relative expression of SlETR6 under high salt stress
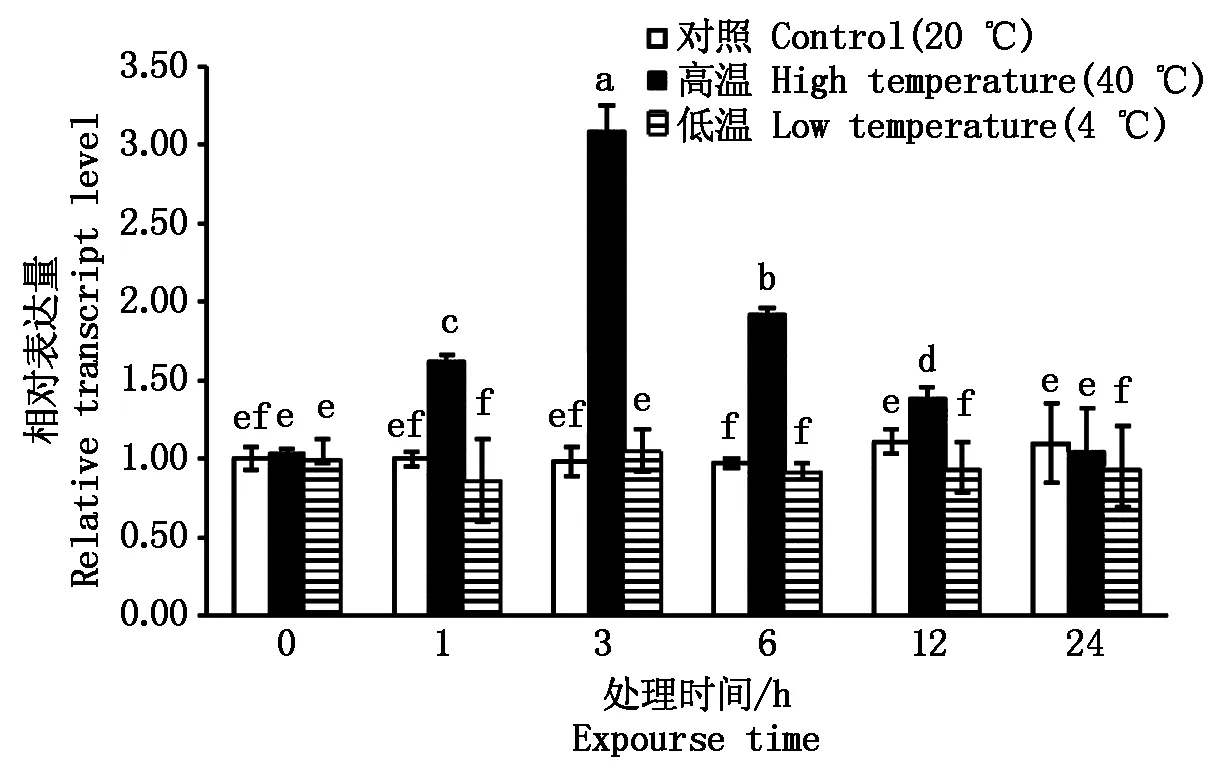
圖6 溫度脅迫下SlETR6基因的表達情況Fig.6 Relative expression of SlETR6 under temperature stress
2.6 SlETR6基因在干旱脅迫下的表達分析
在干旱脅迫條件下,SlETR6基因脅迫處理1 h后迅速應答,與對照組相比,其相對表達量提高1倍,3 h后其表達量呈下降趨勢,但仍高于同期對照組。干旱脅迫處理24 h后,SlETR6基因的相對表達量恢復正常水平(圖7)。與同期對照組相比,SlETR6基因在干旱脅迫的初期(1,3,6 h)均表現出顯著性上調表達。

圖7 干旱脅迫下SlETR6基因的表達情況Fig.7 Relative expression of SlETR6 under drought stress
3 結論與討論
植物抗逆效應是植物對抗不良環境,比如抗旱、抗鹽堿、抗澇、抗風、抗凍、抗病蟲害等的能力。逆境的種類多種多樣,主要可分為生物逆境和非生物逆境兩大類。研究植物在非生物脅迫條件下的調節機制,對通過植物育種改變作物的特性、培育抗逆新品種有重要的意義。乙烯在響應植物逆境脅迫過程中有重要的作用[25-29],而對于乙烯受體在響應非生物脅迫過程中的研究較少,SlETR6是番茄乙烯受體家族成員之一,目前,尚無SlETR6參與非生物脅迫應答中的功能研究相關報道。
本研究在番茄中克隆了乙烯受體家族成員之一SlETR6,含有2 265 bp的開放讀碼框,編碼754個氨基酸。乙烯受體是乙烯信號轉導的負調控因子,參與了植物生長發育的多個過程。序列分析及同源性比對分析表明,SlETR6與馬鈴薯ETR-like蛋白的同源性最高。啟動子序列分析表明,SlETR6具有多種逆境響應功能元件,如熱脅迫、干旱脅迫、低氧脅迫、光響應、激素(乙烯、水楊酸、赤霉素)誘導響應、防御與脅迫響應元件等。
通過實時熒光定量PCR分析了SlETR6基因在種子、根、葉、花、花后5,10,15,25,35(綠熟期果實),45 d(紅熟期果實)等不同組織中的相對表達量,SlETR6在番茄的不同組織中均有表達,且在花和轉色期果實中表達量較高,推測SlETR6可能對于花的發育及果實的成熟過程有一定的調控作用。
另外,SlETR6對不同脅迫的響應模式不同。在4,40 ℃ 2個溫度逆境脅迫環境下,SlETR6受到高溫脅迫后強烈誘導,且在處理后3 h左右達到峰值,隨后表達水平小幅下調,但仍然顯著高于對照組,在處理后24 h恢復正常水平;高鹽脅迫下,SlETR6在處理6 h后被誘導呈上調表達,并在處理后12 h達到峰值,為對照組的3.6倍;SlETR6在干旱脅迫早期受到強烈誘導,在處理后1 h達到峰值,是對照組的2倍。qPCR結果表明SlETR6同時受到高鹽、高溫、干旱等脅迫條件誘導,且生物信息學分析表明SlETR6基因啟動子序列含有大量脅迫應答元件,包括干旱、熱、防御反應相關的元件。綜上,推測SlETR6基因在響應脅迫應答中發揮重要調控作用。本研究為后續研究SlETR6在響應逆境脅迫過程中的功能及作用機制奠定基礎,為將來利用分子育種手段培育番茄抗逆新品種提供重要的基因資源。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
