2 種天然抗氧化劑與鰱魚肌球蛋白的相互作用
2019-03-11 08:44:00岳世陽熊善柏杜紅英
食品科學 2019年4期
黃 淵,岳世陽,熊善柏,2,杜紅英,2,*
(1.華中農業大學食品科學技術學院,湖北 武漢 430070;2.國家大宗淡水魚加工技術研發分中心(武漢),湖北 武漢 430070)
我國是淡水魚類產品生產大國,2016年全國淡水產品總產量3 411.11萬 t,其中魚類產量2 986.65萬 t,占全年淡水產品總量的87.56%[1]。但是由于魚類產品水分和蛋白質含量高,且自身攜帶較多的細菌,在貯藏、運輸和銷售過程中,容易失去其原有鮮味,造成腐敗變質。目前魚類的保鮮主要是冷藏為主,輔以添加防腐劑或抗氧化劑,但一些合成的防腐劑或抗氧化劑存在一定的致癌風險[2],不符合現在的生產消費要求,所以加大開發健康天然的防腐劑或抗氧化劑成為魚類保鮮的研究重點。
表沒食子兒茶素沒食子酸酯(epigallocatechin gallate,EGCG)(圖1A)是從中國綠茶中提取的一種成分,是綠茶兒茶素中含量最高的組分,約占綠茶兒茶素的50%~80%,綠茶所表現的很多生物活性主要來源于EGCG[3]。李鵬等[4]通過測定不同濃度EGCG處理的羅非魚片新鮮度的變化,證明EGCG可有效提高冷藏期間羅非魚片的品質,延長其貨架期,且0.2%的EGCG對羅非魚片的保鮮效果更佳。由于EGCG易氧化,影響其保鮮效果,所以駱曉波[5]以納米脂質體為載體,制成EGCG納米脂質體并研究其穩定性以及對鱈魚的保鮮效果,證明EGCG納米脂質體在維持pH值的穩定,防止魚肉氧化、腐敗變質,維持感官性狀,抑制細菌的生長等方面都表現出優勢,且無毒副作用,適合于魚肉的保鮮。
白藜蘆醇(resveratrol,RE)(圖1B),化學名稱為3,4,5-三羥基二苯乙烯,是含有芪類結構的酚化合物,主要存在于葡萄、虎杖、花生、桑椹等植物中,尤其在種皮中含量較高[6]。有研究表明,RE對月季切花有一定的保鮮作用[7],有效調節切花水分平衡,減輕細胞質膜透性,降低膜脂過氧化水平,從而延緩切花衰老進程,延長瓶插壽命;RE處理對碭山酥梨有較好的保鮮效果[8],可抑制共軛三烯的含量、增加細胞的膜透性、降低α-法尼烯的峰值,顯著抑制和延緩梨果實在貯藏期和貨架期黑皮病的發生。將RE應用于植物以及植物制品保鮮中的研究國內外均有報導[8-9],但關于RE對魚類保鮮效果及其與肌球蛋白作用的研究鮮見報道。因此,本實驗擬就其與肌球蛋白相互作用方面進行研究。
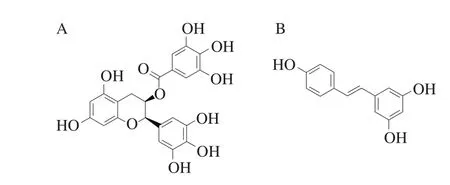
圖1 EGCG(A)、RE(B)結構式Fig. 1 Chemical structures of EGCG (A) and RE (B)
蛋白質為肌肉組織的主要組成部分,其降解、聚合和變性都會導致蛋白質功能的喪失,從而直接決定魚類肌肉質量的好壞以及魚類腐敗變質[10]。其中肌球蛋白是各類肌細胞中含量最多的結構蛋白和收縮蛋白,是構成肌肉肌原纖維粗絲的基本組成蛋白,是橫紋肌細胞表達最豐富的蛋白,約占其總蛋白庫的25%[11]。本實驗以EGCG、RE和肌球蛋白為原料,運用熒光光譜、圓二色譜研究EGCG、RE與肌球蛋白的相互作用,由熒光猝滅現象和Forster非輻射能量轉移理論求得EGCG、RE與肌球蛋白的結合常數和結合位點數,從熱力學參數推出二者之間的作用力類型,并探討EGCG、RE對肌球蛋白構象的影響,以期為EGCG、RE在魚類保鮮應用提供理論依據。
1 材料與方法
1.1 材料與試劑
鰱魚(體質量2~3 kg) 市購;EGCG、RE(均為色譜純,純度≥98%) 上海源葉生物科技有限公司;其他試劑均為分析純,實驗用水均為二次去離子水。
1.2 儀器與設備
F-4500型熒光分光光度計 日本日立公司;AE-240雙量程電子分析天平 瑞士Metter-Toledo公司;J-1500型圓二色譜儀 日本分光株式會社;TB-85恒溫水浴裝置 日本島津公司。
1.3 方法
1.3.1 提取肌球蛋白
參考Park等[12]的方法,并略作修改。尸僵前的鰱魚背脊肉用食品調理機破碎,向破碎后的肌肉中加入10 倍體積的溶液A(0.1 mol/L KCl、0.02% NaN3、20 mmol/L Tris-HCl,pH 7.5)。并用高速分散均質機于9 000 r/min條件下均質1 min,均質液于4 ℃條件下放置15 min,5 000 r/min離心5 min,沉淀于5 倍體積溶液B(0.45 mol/L KCl、5 mmol/L β-巰基乙醇、0.2 mol/L醋酸鎂、1 mmol/L乙二醇-雙-(2-氨基乙基)四乙酸、20 mmol/L Tris-HCl,pH 6.8)中溶解,同時加入ATP-Na2使肌球蛋白與肌動蛋白解離,魚肉肌球蛋白的提取需加ATP-Na2至終濃度為5 mmol/L,于4 ℃條件下放置1 h后4 ℃、10 000 r/min離心10 min,上清液用10 倍體積1 mmol/L KHCO3溶液稀釋,于4 ℃條件下放置1 h,然后4 ℃、12 000 r/min離心10 min;沉淀重新用2.5 倍體積溶液C(0.5 mol/L KCl、5 mmol/L β-巰基乙醇、20 mmol/L Tris-HCl緩沖液,pH 7.5)溶解,于4 ℃條件下放置10 min,再用2.5 倍體積的1 mmol/L KHCO3溶液稀釋并加MgCl2至終濃度為10 mmol/L,于4 ℃條件下放置過夜,然后10 000 r/min離心10 min;沉淀物用0.5 倍體積溶液D溶解(0.6 mol/L NaCl、20 mmol/L Tris-HCl緩沖液,pH 7.5),即為肌球蛋白溶液。采用福林-酚法[13]測定蛋白濃度。
1.3.2 熒光光譜測定
分別移取一定量的EGCG和RE乙醇溶液于10 mL具塞比色管中,再加入1 mg/mL肌球蛋白溶液1.0 mL,用0.6 mol/L NaCl(20 mmol/L Tris-HCl,pH 7.5)緩沖溶液稀釋,多酚濃度[14]分別為0、1×10-5、2×10-5、3×10-5、4×10-5mol/L,在實驗溫度條件下恒溫1 h。以280 nm為激發波長,狹縫寬5 nm,掃描300~450 nm波長范圍內的熒光光譜。設置波長差(Δλ)分別為15、60 nm,狹縫寬均為5 nm,掃描速率240 nm/min,掃描260~380 nm范圍內的同步熒光光譜[15]。
1.3.3 圓二色譜測定
試劑處理方式同1.3.2節,設置波長掃描范圍190~250 nm,掃描速率50 nm/min,分辨0.1 nm,響應時間1 s,石英品池光徑1 mm,N2流量4 L/min[16]。
1.3.4 表面疏水性測定
采用8-苯氨基-1-萘磺酸(1-anilino naphthalene-8-sulfonic acid,ANS)熒光探針法測量蛋白質表面疏水性[17]。將已知蛋白分別稀釋為0、0.05、0.1、0.15、0.2 mg/mL,同時加入多酚,使EGCG和RE濃度均分別為0、1×10-5、2×10-5、3×10-5、4×10-5mol/L。每4 mL溶液加入20 μL 8×10-3mol/L ANS-0.1 mol/L磷酸鹽緩沖液(pH 7.4)。反應10 min后測其熒光強度。設置狹縫寬度5 nm,激發波長390 nm,發射波長479 nm。表面疏水性以熒光值-蛋白濃度曲線的斜率來表示。
1.4 數據處理
采用SAS 9.2軟件進行數據分析,運用Origin 9.0軟件進行圖形繪制及線性擬合。
2 結果與分析
2.1 熒光猝滅光譜
肌球蛋白分子中因含有在激發波長為280 nm條件下能發射熒光的色氨酸、酪氨酸等氨基酸殘基而具有內源性熒光,其中色氨酸殘基是主要的熒光貢獻基團[18]。當激發波長為280 nm時,由圖2A可知,肌球蛋白在波長341 nm處具有強熒光發射峰,且隨著EGCG濃度的增加,肌球蛋白的內源性熒光強度有規律地降低,表明EGCG可以大幅度猝滅肌球蛋白內源性熒光,這說明EGCG、肌球蛋白發生了相互作用;由圖2B可知,隨著RE濃度的增加,肌球蛋白內源性熒光也呈現出大幅度有規律的降低,且伴隨著明顯的熒光發射峰紅移現象(從341 nm到365 nm),說明RE與肌球蛋白發生了相互作用,使其發色團微環境空間構象發生變化,即色氨酸殘基所處環境極性增強,疏水性減弱。
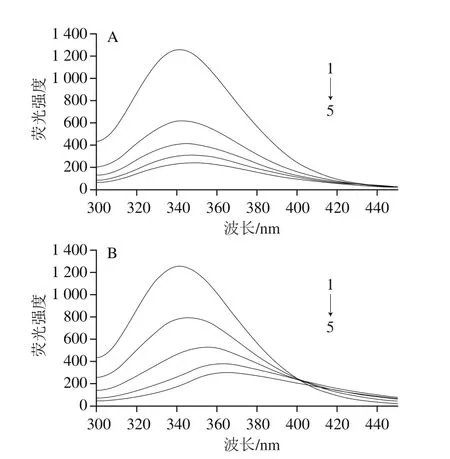
圖2 EGCG(A)和RE(B)對肌球蛋白的猝滅作用Fig. 2 Fluorescence quenching spectra of myosin with EGCG (A) and RE (B)
2.2 熒光猝滅機理
溶液中熒光的猝滅,指的是具備內源性熒光物質分子與溶劑分子之間所發生的導致熒光強度下降的物理或化學的作用過程。熒光物質分子同猝滅劑相互作用方式的差異產生了不同的猝滅方式,例如動態猝滅、靜態猝滅等形式。動態猝滅,是指猝滅劑與處于激發態的熒光物質分子之間發生相互作用而引起的一種猝滅過程;靜態猝滅,是指猝滅劑與處于基態的熒光物質分子發生相互作用所引起的一種猝滅過程[19]。其中,動態猝滅過程符合Stern-Volmer方程[20],如方程(1)所示:
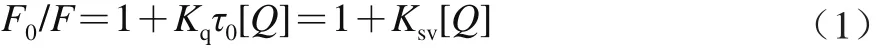
式中:F0為未加入猝滅劑時的熒光強度;F為加入猝滅劑后的熒光強度;Kq為雙分子猝滅過程速率常數/(L/(mol·s));Ksv為Stern-Volmer動態猝滅常數/(L/mol);[Q]為猝滅劑的濃度/(mol/L);τ0為猝滅劑不存在條件下生物大分子的平均熒光壽命(約為10-8s)。
為確定EGCG、RE與肌球蛋白相互作用的猝滅機理,先用Stern-Volmer方程處理數據[21],分別得到不同溫度條件下EGCG、RE與肌球蛋白熒光猝滅的Stern-Volmer方程,如圖3所示。
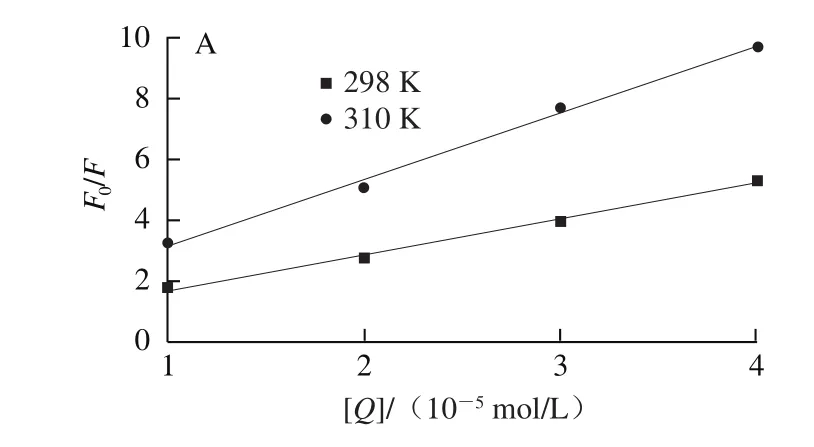

圖3 不同溫度條件下EGCG(A)和RE(B)對肌球蛋白熒光猝滅的Stern-Volmer曲線Fig. 3 Stern-Volmer plots for fl uorescence quenching of EGCG (A)-myosin and RE (B)-myosin systems at different temperatures

表1 不同溫度條件下EGCG和RE與肌球蛋白分別相互作用的Stern-Volmer猝滅常數KsvTable 1 Stern-Volmer quenching constants for EGCG-myosin and RE-myosin systems at different temperatures
對圖3進行數學計算可求得EGCG和RE與肌球蛋白相互作用的Ksv和Kq,如表1所示。由圖3A可知,EGCG與肌球蛋白的Stern-Volmer猝滅曲線中的F0/F和[Q]之間存在線性相關性,且斜率隨著溫度的升高而增加,說明EGCG與肌球蛋白之間發生了動態猝滅現象,同時不同溫度曲線條件下的Kq值均大于最大動態猝滅速率2.0×1010L/(mol·s),說明EGCG與肌球蛋白的相互作用過程中也存在靜態猝滅的發生,所以EGCG與肌球蛋白的相互作用過程既有動態猝滅也有靜態猝滅[22]。由圖3B可以看出,RE與肌球蛋白的Stern-Volmer猝滅曲線中的F0/F和[Q]之間也存在線性相關性,但是斜率是隨著溫度的升高而降低,且不同溫度條件下Kq值也都大于最大動態猝滅速率2.0×1010L/(mol·s),結合其熒光光譜圖出現紅移現象,說明RE與肌球蛋白的相互作用過程不存在動態猝滅而是形成復合物的靜態猝滅[23]。
2.3 結合常數和結合位點數
在靜態猝滅過程中,熒光物質與猝滅劑分子之間的結合常數可以通過靜態猝滅公式(2)[24]計算:
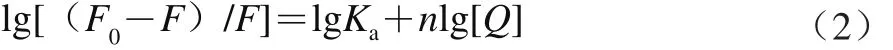
式中:F0為未加入猝滅劑時的熒光強度;F為加入猝滅劑后的熒光強度;Ka為結合常數/(L/mol);[Q]為配合物的濃度/(mol/L);n為結合位點數。
得到lg[(F0-F)/F]與lg[Q]的關系和相關方程,如圖4所示。圖4中擬合曲線的斜率代表多酚與肌球蛋白的結合位點數n值;擬合曲線的截距代表化合物與肌球蛋白的結合常數Ka值。
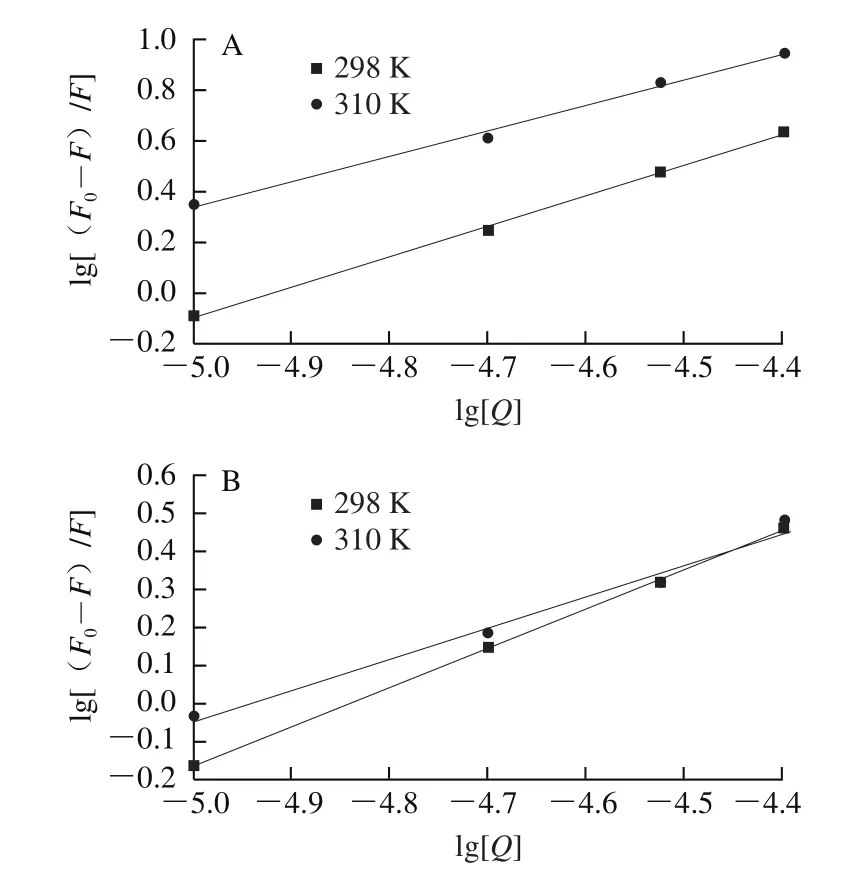
圖4 不同溫度條件下EGCG(A)和RE(B)與肌球蛋白的lg[(F0-F)/F]與lg[Q]的關系圖Fig. 4 Plot of lg[(F0-F)/F] versus lg[Q] for EGCG (A)-myosin and RE (B)-myosin systems at different temperatures
對圖4進行觀察可得EGCG、RE與肌球蛋白反應的結合常數和結合位點數,如表2所示。EGCG和RE與肌球蛋白相互作用的結合常數、結合位點數都隨著溫度上升而降低,表現出明顯的靜態猝滅,并且多酚與肌球蛋白的結合位點數均接近1,表明多酚與肌球蛋白的之間結合力很好。

表2 不同溫度條件下EGCG和RE分別與肌球蛋白結合的相關常數和方程Table 2 Linear regression equations and correlation coef ficients for EGCG-myosin and RE-myosin interactions at different temperatures
2.4 多酚與肌球蛋白之間作用力類型
有機小分子和蛋白質之間的作用力主要有疏水作用力、氫鍵、范德華力和靜電引力等。不同小分子與蛋白質結合力類型不同。Ross等[25]根據大量的實驗結果,總結出根據反應前后熱力學焓變ΔH和熵變ΔS的相對大小,判斷藥物與蛋白質之間的主要作用力類型的規律。ΔH大于0、ΔS大于0為疏水作用力;ΔS小于0、ΔH小于0為氫鍵和范德華力;ΔH小于0、ΔS大于0為靜電引力。在溫度變化不大時,反應的焓變ΔH可以看作一個常數,由結合常數Ka,按公式(3)求出反應的自由能變(ΔG),然后根據公式(4)、(5)分別求出ΔH、ΔS,如表3所示。

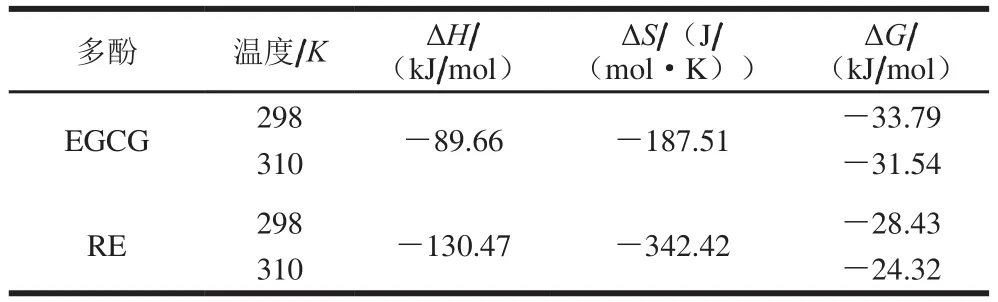
表3 EGCG、RE與肌球蛋白結合的熱力學參數Table 3 Thermodynamic parameters of EGCG-myosin and RE-myosin interactions
由表3可知,EGCG與肌球蛋白作用過程中的ΔH小于0、ΔS小于0,說明EGCG與肌球蛋白之間的主要作用為氧鍵和范德華力。而ΔG小于0,表明EGCG與肌球蛋白的結合為自發過程。同樣,RE與肌球蛋白作用過程中的ΔH小于0、ΔS小于0,表明了RE與肌球蛋白之間的主要作用力也為氫鍵和范德華力。而ΔG小于0,表明RE與肌球蛋白的結合也為自發過程。
2.5 同步熒光光譜
同步熒光光譜因其圖譜簡單、譜帶窄、能夠減少光譜重疊等優點,是用來研究小分子與蛋白質相互作用對蛋白質發色團分子微環境構象改變的好方法[26]。當設置Δλ為15 nm時,只顯示酪氨酸殘基的熒光特征光譜,當設置Δλ為60 nm時,只顯示色氨酸殘基的熒光特征光譜[27]。因此由最大發射波長的改變可判斷蛋白質酪氨酸殘基和色氨酸殘基微環境的極性變化,當最大吸收波長藍移時,說明多酚的加入使環境疏水性增加,當最大吸收波長紅移時,說明環境疏水性減小,蛋白分子趨向于伸展[28]。
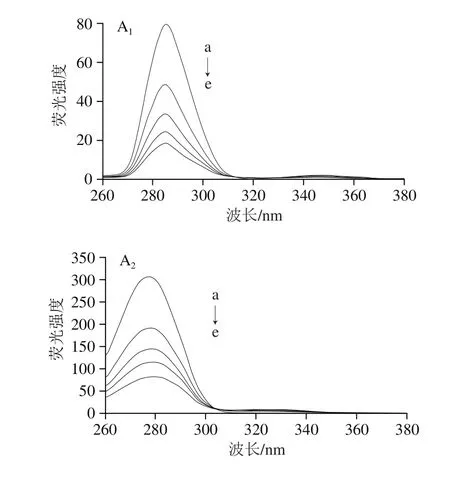
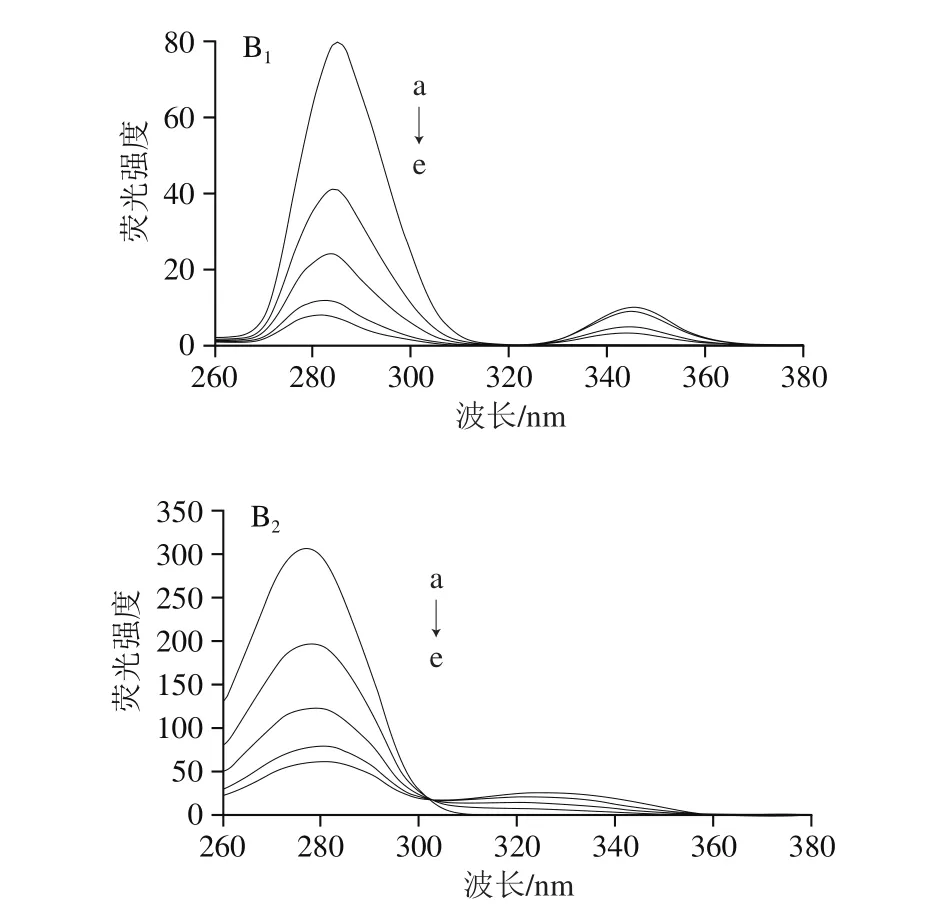
圖5 EGCG(A)和RE(B)與肌球蛋白相互作用的同步熒光光譜Fig. 5 Synchronous fl uorescence spectra of EGCG (A)-myosin and RE (B)-myosin systems
從圖5A可以看出,當Δλ為15 nm時,隨著EGCG濃度的增加,酪氨酸殘基的熒光強度明顯下降,但最大吸收波長沒有變化,當Δλ為60 nm時,隨著EGCG濃度的增加,色氨酸殘基的熒光強度也明顯下降,且伴有輕微紅移,說明EGCG與肌球蛋白相互作用均會導致酪氨酸殘基和色氨酸殘基的熒光猝滅,并且色氨酸殘基的熒光貢獻比酪氨酸多,這與激發波長的設置以及色氨酸和酪氨酸的相對含量有關[29],而且EGCG影響了色氨酸殘基的微環境,使該區域極性增加,疏水性降低[28]。
從圖5B可以看出,當Δλ為15 nm時,隨著RE的加入,酪氨酸殘基的熒光強度明顯下降,且伴有明顯藍移,說明RE改變了酪氨酸殘基的微環境,使該區域極性減小,疏水性增大;當Δλ為60 nm時,隨著RE的加入,色氨酸殘基的熒光強度也明顯下降,且伴有輕微紅移,說明RE也改變了肌球蛋白色氨酸殘基的微環境,使該區域極性增大,疏水性降低。
2.6 圓二色譜測定結果
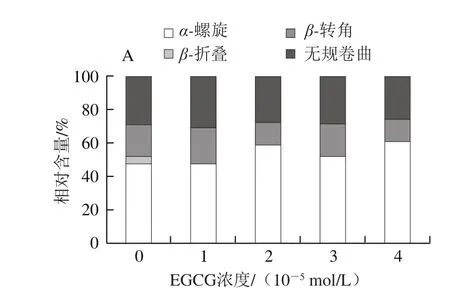
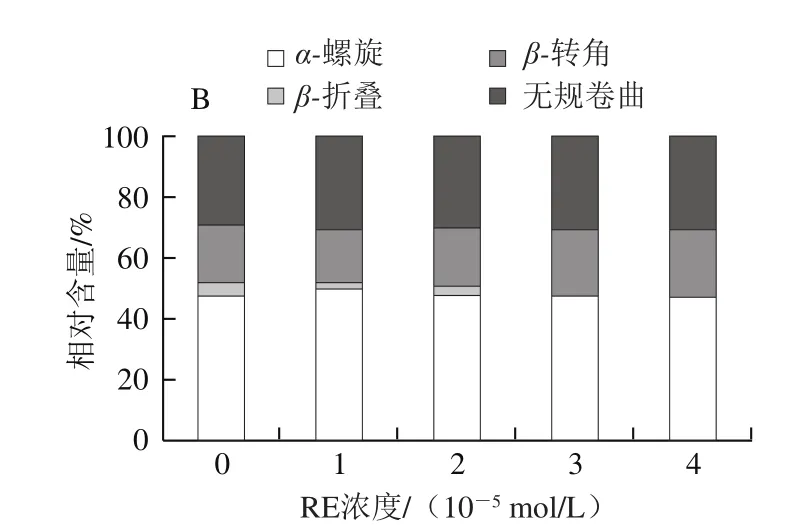
圖6 EGCG(A)、RE(B)與肌球蛋白相互作用的圓二色譜圖Fig. 6 CD spectra of EGCG (A)-myosin and RE (B)-myosin systems
從圖6可以看出,隨著EGCG濃度上升,肌球蛋白α-螺旋相對含量整體呈上升趨勢,β-折疊、β-轉角、無規卷曲都呈現下降趨勢,說明EGCG與肌球蛋白作用使肌球蛋白二級結構發生變化,肽鏈收縮;而隨著RE濃度上升,肌球蛋白α-螺旋、無規卷曲含量基本不變,僅β-轉角、β-折疊含量產生少許變化,說明RE對肌球蛋白二級結構沒明顯影響。
2.7 表面疏水性測定結果
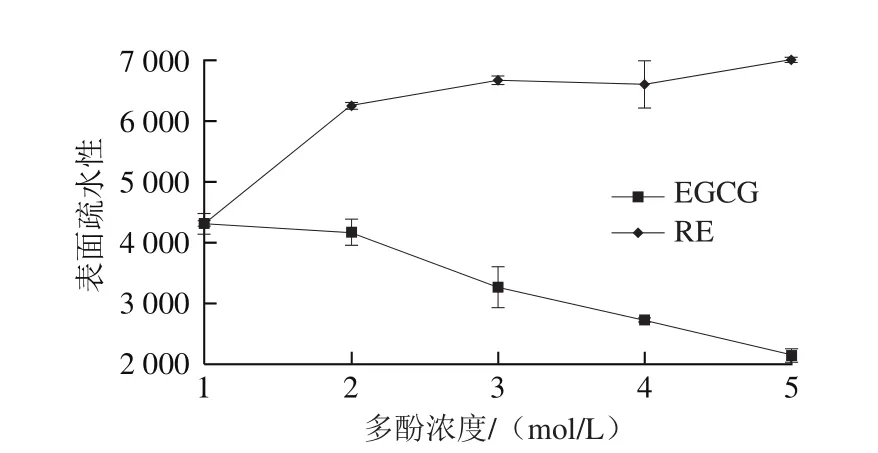
圖7 EGCG、RE對肌球蛋白表面疏水性影響Fig. 7 Effects of EGCG and RE on surface hydrophobicity of myosin
蛋白質作為具有復雜空間結構的生物大分子,其構象的變化必然會影響其表面氨基酸的分布、密度以及疏水空穴的數量,而使表面疏水性發生變化[30]。為進一步說明EGCG、RE對肌球蛋白理化性質的影響,考察EGCG、RE對肌球蛋表面疏水性的影響情況。從圖7可看出,隨著EGCG濃度上升,肌球蛋白表面疏水性呈下降趨勢,這與同步熒光數據得出的EGCG使色氨酸殘基微環境疏水性減少和圓二色譜數據得出的EGCG使α-螺旋增加的結果相匹配;隨著RE濃度上升,肌球蛋白表面疏水性呈上升趨勢,也與同步熒光數據有一定的相關性[31]。
3 結 論
EGCG、RE對肌球蛋白的內源熒光都具有較強猝滅作用,這種猝滅主要為靜態猝滅。求得EGCG、RE與肌球蛋白的結合常數、結合位點和熱力學參數,且熱力學數據(ΔH<0、ΔS<0、ΔG<0)表明它們主要靠氫鍵和范德華力結合并且是自發過程;同時用同步熒光光譜、圓二色譜和表面疏水性測定探討EGCG、RE對肌球蛋白構象的影響,結果表明EGCG會引起肌球蛋白二級結構變化,α-螺旋相對含量增加,降低肌球蛋白表面疏水性,RE對肌球蛋白二級結構無明顯影響,使其表面疏水性增加。
