曼氏無針烏賊染色體核型分析
2019-03-25 01:10:28高天翔王露萍遲長鳳江麗華王世鋒
浙江海洋大學學報(自然科學版) 2019年4期
高天翔,王露萍,遲長鳳,江麗華,王世鋒
(1.浙江海洋大學水產學院,浙江舟山 316022;2.國家海洋設施養殖工程技術研究中心,浙江舟山 316022;3.浙江海洋大學海洋科學與技術學院,浙江舟山 316022;4.海南大學海洋學院,海南海口 570228)
曼氏無針烏賊(Sepiella maindroni de Rochebrune)俗稱烏賊、墨魚、血墨。分布于渤海、黃海、東海、南海,為印度洋和西太平洋廣分布種,其中心分布區在中國浙江近海和閩東海域。曼氏無針烏賊是東海重要經濟種類,20 世紀50-70 年代曾是舟山漁場“四大海產”之一,但由于資源的不合理利用,以及生態環境的破壞,80 年代中后期,浙江沿海漁場曼氏無針烏賊產量急劇下降,已形不成漁汛,瀕臨滅絕[1]。21 世紀初,浙江海洋學院和寧波大學先后開展了曼氏無針烏賊人工繁育技術研究,并取得突破。2007-2009 年浙江海洋學院研究團隊在浙江沿海的舟山東極海域、臺州大陳海域、溫州洞頭海域開展小規模曼氏無針烏賊苗種(受精卵)探索性增殖放流研究[2]。近年來,福建省和江蘇省也相繼開展了曼氏無針烏賊的增殖放流工作。隨著人工繁育技術的突破和人工增殖放流工作的開展,目前資源量得到了顯著恢復,據相關報道資源量已恢復至2 000 t 的水平[3]。目前有關曼氏無針烏賊的研究工作主要集中在繁殖生物學[4-6]、增殖放流[1,7]及營養飼料[8-10]等方面。
染色體是生物發育、進化、遺傳和變異的物質基礎,研究染色體的行為、數目、核型和帶型等對于了解生物的遺傳組成、遺傳變異規律、發育機制、物種起源和親緣關系、進化地位、分類及種族關系等都具有重要的參考價值[11]。目前有關頭足類染色體的研究國內外已開展較多研究:萊氏擬烏賊Sepioteuthis lessoniana[12]、長蛸Octopus minor[13]、金烏賊Sepia esculenta[14]、鸚鵡螺Nautilus macromphalus[15]等。但有關曼氏無針烏賊染色體核型分析的研究在國內外尚未見報道。本研究制備了曼氏無針烏賊染色體并進行了核型分析,以期為曼氏無針烏賊的遺傳背景和資源保護提供種質特征方面的相關資料。
1 材料與方法
1.1 實驗材料
曼氏無針烏賊親體為舟山漁場的野生烏賊,選擇體質健康、性成熟個體,在室內人工養殖,實驗樣品取自親體產出的受精卵和不同發育階段的胚胎。
1.2 實驗方法
1.2.1 染色體標本的制備
參照王曉華等[14]的金烏賊染色體制備方法,并對其方法進行優化。選取紅眼期曼氏無針烏賊胚胎,剝去卵膜,取出胚胎。將胚胎浸入0.04%的秋水仙素溶液,浸泡處理1 h,去凈秋水仙素,用0.075 mol·L-1KCl低滲處理45 min,離心,移去低滲液,加入4℃預冷的卡諾氏國定液(甲醇:冰醋酸=3:1)固定,間隔15 min換固定液1 次,連續固定3 次,然后加入50%冰醋酸進行解離,解離時間為45 min。將解離好的樣品熱滴法滴片,空氣干燥過夜。
1.2.2 染色體數目統計及核型分析
制備好的染色體玻片用10% Giemsa 染液染色20 min,蒸餾水沖洗,自然晾干后進行顯微觀察和拍照。選擇分散良好、形態清晰、兩條臂適當分開且數目完整的中期分裂相約40 個進行拍照。進行染色體計數、測量,并根據LEVAN A,et al[16]的分類標準進行染色體配對、分組,制作核型圖。
2 結果
2.1 染色體數目
實驗對40 個分散良好的中期分裂相進行染色體計數,曼氏無針烏賊二倍體染色體數為92 的約占所統計中期分裂相的91%,占絕對優勢,由此可以判定曼氏無針烏賊的二倍體染色體數目為2n=92。
2.2 染色體核型
選取3 個分散良好,形態清晰的中期分裂相進行測量和統計分析,獲得曼氏無針烏賊染色體的相對長度[染色體相對長度=(單條染色體長度×2/全部染色體總長度)×100]、臂比及其核型,如圖1 和表1 所示。根據LEVAN,et al[16]提出的核型分類標準,可將曼氏無針烏賊染色體分為4 個類型,其中,m 型染色體28 對,sm 型12 對,st 型4 對,t 型3對,核型公式為2n=54m+24sm+8st+6t,臂數(NF)=170。

圖1 曼氏無針烏賊染色體中期分裂相(A)及核型(B)Fig.1 Metaphase chromosomes (A) and karyotype (B) of S.maindroni
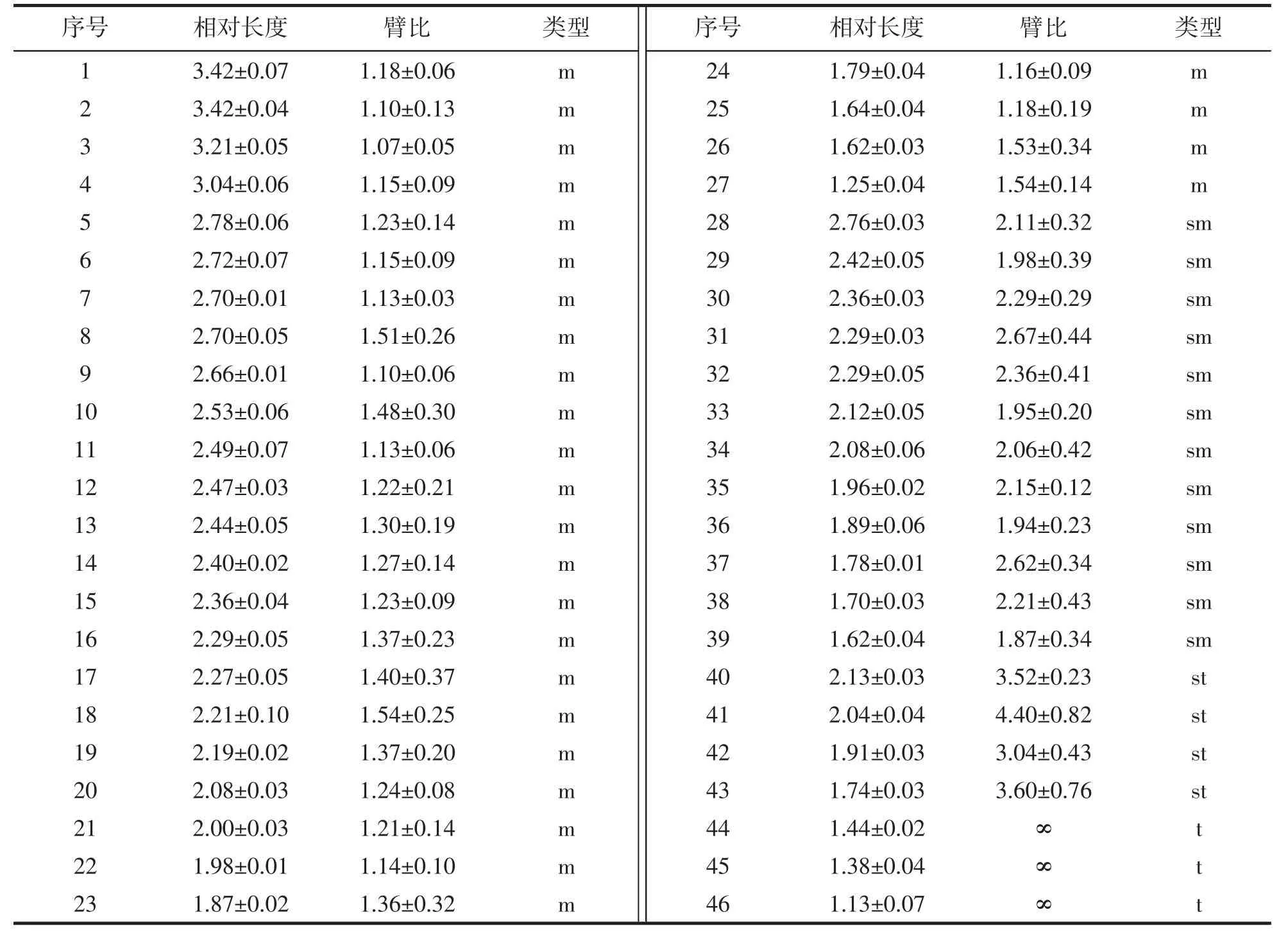
表1 曼氏無針烏賊染色體相對長度及臂比值(平均值±標準差)Tab.1 Relative length and ratio of chromosomes in S.maindroni
3 討論
3.1 染色體的制備
提取到新陳代謝旺盛、分裂增生較快的組織是成功制備染色體的基本條件[17-18]。魚類染色體的獲得通常采用體內注射PHA 腎細胞培養法[18],但是由于頭足類染色體數目多及形態大小適中的染色體中期分裂相較難獲得等原因。據報道,用于頭足類染色體的制備組織有3 種:(1)雄性的精細胞[19-20];(2)鰓心血液[17];(3)胚胎[12-13,21]。采用胚胎制作頭足類染色體可以獲得比較理想的結果[12]。本研究在制備曼氏無針烏賊染色體的過程中,分別選取了受精卵和不同發育時期的胚胎作為染色體制備材料。實驗發現只有以紅眼期的曼氏無針烏賊胚胎為材料制備的染色體中,染色體分散最好,中期分裂相對較多,且染色體的形態最為清晰。因此,本研究認為曼氏無針烏賊染色體標本的制備材料以紅眼期胚胎最佳。
3.2 染色體核型與種質鑒定關系
染色體是遺傳物質的主要載體,染色體核型研究是細胞遺傳學的基礎,染色體組型在很大程度上具有物種的特異性,特定的生物體具有特定數目、形態特征的染色體[22-23]。染色體數目和結構對于研究物種進化具有重要作用。目前國內外頭足類染色體核型研究僅13 例(表2)。GAO Yuemian,et al[12]成功對5 種頭足類染色體核型進行了描述,這5 種頭足類染色體數目均為92 條,核型稍有差別。而在此之前的很多研究,由于未獲得好的中期分裂相或者顯微拍照質量等原因,僅進行了染色體數目描述。WANG Jinhai,et al[13]對3 種蛸類做了染色體核型分析,發現基于染色體核型進化距離和核型近似系數的頭足類進化分析,與基于線粒體全序列的頭足類進化樹結果一致,可以作為傳統分類學和分子系統分析的補充,有望成為常規系統發育的輔助手段。然而在魚類研究中,由于魚類核型基本都是端部著絲粒染色體,核型進化距離和近似系數法并不適用。已有的頭足類染色體研究結果與基于線粒體細胞色素氧化酶亞基I 的建樹結果相一致(圖3),其中鸚鵡螺染色體數目為52 條,是頭足類最為古老的物種,而八腕總目染色體數目均為60 條,十腕總目染色體數除阿拉伯烏賊染色體數目為68 條外,其余均為92 條。十腕總目染色體數目具有高度一致性,但是不同種類的核型不同,可作為鑒別不同種屬的重要依據。目前,頭足類分類最傳統的方法是依據其形態特征,通過對個體的分節特征、度量特征和解剖學特征的測量和鑒定,依據這些特征的差異進行分類[24]。但是頭足類在受外界刺激或死亡狀態下體色容易改變;另外,大多數頭足類的幼體階段外部特征比較相似。這些問題都為僅僅依靠形態學的方法鑒別頭足類種類帶來困難。因此,在頭足類種類鑒別過程中,無法通過形態特征進行區分時,可以考慮使用染色體核型的方法。染色體核型的多樣化為頭足類的系統分類和種質鑒定有著重要的輔助作用。
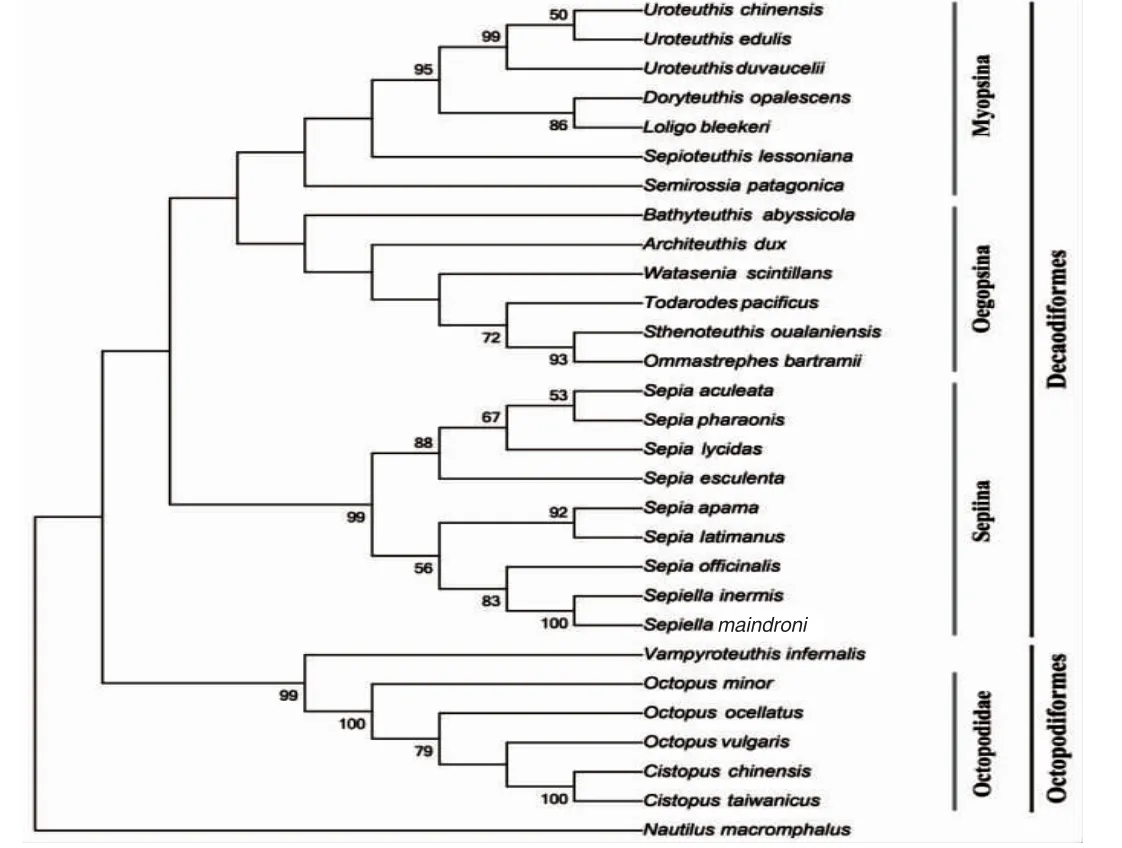
圖3 基于線粒體細胞色素氧化酶亞基I(COI)的頭足類進化樹Fig.3 Phylogenetic relationships among the cephalopods based on mitochondrial COI gene sequence
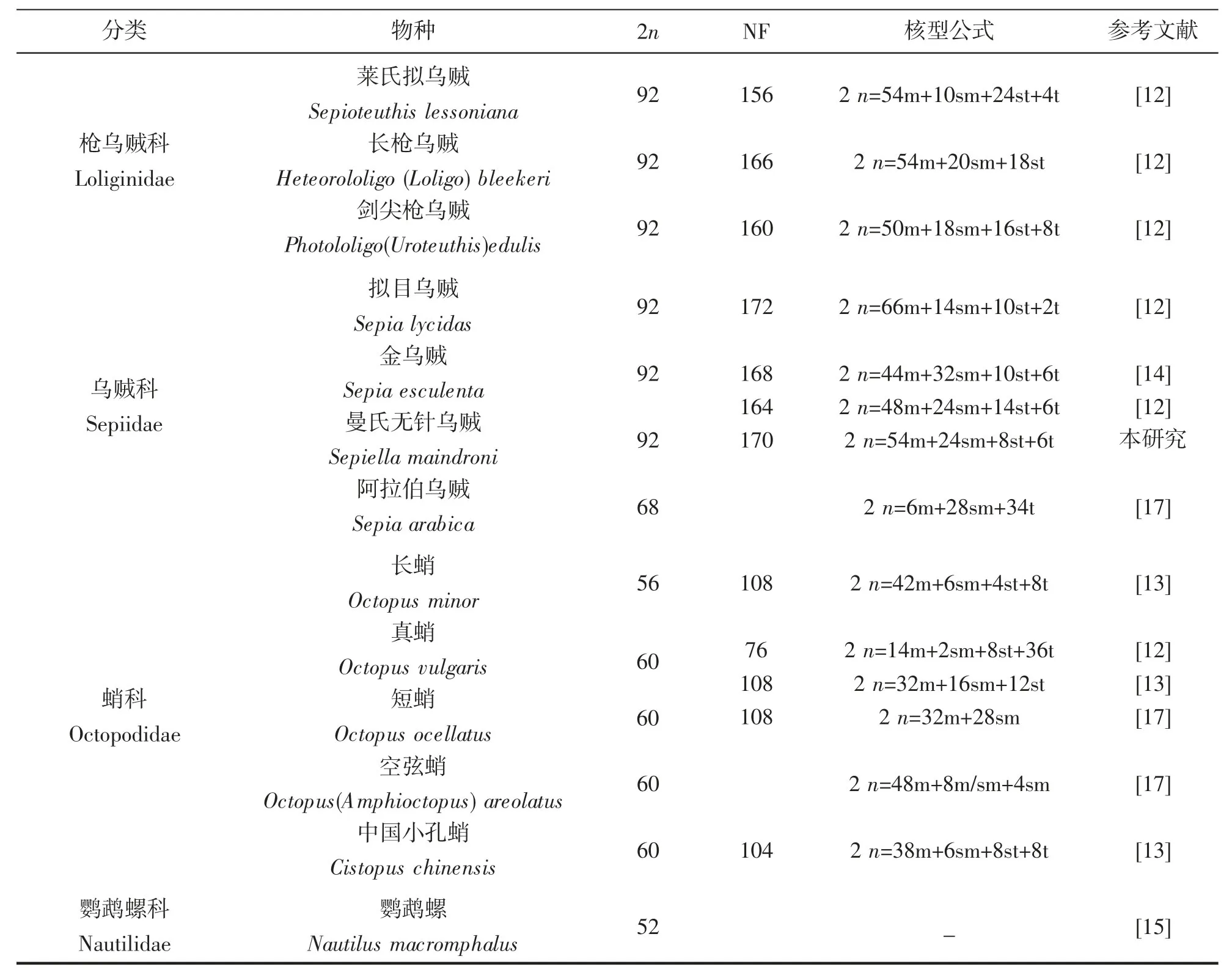
表2 部分已報道頭足類染色體核型Tab.2 Karyotypes of several reported species of the cephalopods
為了更好研究頭足類染色體遺傳信息,在今后研究中應采用除染色體核型以外,開展帶型研究、染色體核仁組織區及熒光原位雜交技術的綜合分析。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
考試與評價·高二版(2021年5期)2021-09-10 07:22:44
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
非公有制企業黨建(2020年9期)2020-09-26 13:22:18
科技傳播(2019年22期)2020-01-14 03:06:54
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
瘋狂英語·新策略(2018年1期)2018-08-20 07:50:38
汽車工程學報(2017年2期)2017-07-05 08:13:02
領導決策信息(2017年12期)2017-05-17 04:49:18
