低氮脅迫對蛋白核小球藻生化組分和絮凝性能的影響
2019-04-09 03:50:30趙艷,汪成
植物營養與肥料學報 2019年3期
關鍵詞:油脂
趙 艷,汪 成
(浙江工商大學食品與生物工程學院,浙江杭州 310018)
以小球藻為代表的微藻是一類能進行光合自養生長的單細胞綠藻,易于培養且生長迅速,富含蛋白、油脂、維生素、礦物質及多種生理活性成分,在醫藥、食品、飼料和生物柴油新能源方面的應用潛力巨大[1-2]。2011年,我國“神舟8號”搭載小球藻上天進行生態循環科學實驗,發現小球藻能提供氧氣,并作為食物來源確保宇航員在空間站的全面營養,而且還可能對很多醫學難題提供重要的解決方案[3]。特別是蛋白核小球藻 (Chlorella pyrenoidosa)具有蛋白含量高,富含各種必需氨基酸和多種營養保健因子,2012年被我國增列為新資源食品[4],其研究開發很快成為熱點。微藻的大規模培養是實現工業化生產的前提。人工培養技術中比基因工程或代謝工程手段更簡單有效的策略是通過改變培養基或培養條件影響藻細胞代謝途徑,以達到提高油脂、多糖等目標化合物生產率的目的[5]。氮是蛋白質、核酸、磷脂和葉綠素等物質合成的基本元素,在新陳代謝中至關重要。研究表明,降低培養基中的氮素供應使藻細胞處于低氮脅迫狀態能影響藻細胞光合效率、生物量與生化組分合成[6]。低氮脅迫作為提高微藻生產效益的低成本技術,增效作用在不同藻種之間差異很大[7],這是因為藻種不同細胞氮代謝途徑和整體代謝網絡模式不同,對低氮脅迫信號的響應機制也不一致。只有針對不同藻種研究低氮脅迫的適宜水平和收獲時間等因素才能使收益最大化。代瑞華等[8]報道適宜的低氮脅迫處理會促進銅綠微囊藻細胞分裂,縮短生長周期。吳桂秀等[9]發現低氮脅迫有利于大真眼點藻和波氏真眼點藻的總脂和中性脂的積累。Adams等[6]研究了低氮脅迫對6種微藻生長與油脂累積的影響,發現普通小球藻經4 mmol/L低氮脅迫培養12天后,其生物量比對照減少了51%,但藻細胞油脂含量增加了302%。Gong等[10]研究了低氮脅迫對4株海洋微藻油脂含量的影響,發現培養5天后角毛藻、三角褐指藻和綠色巴夫藻細胞內總脂肪酸含量已增加了一倍,眼點擬微綠球藻總脂肪酸含量卻增長不明顯。此外,微藻作為一個代謝整體,在氮脅迫下生長和生化組分的合成變化密切相關。如球等鞭金藻 (Isochrysis galbana) 當細胞內源型儲存氮達到最低閾值時,藻細胞生長緩慢同時細胞內的脂質和類胡蘿卜素開始大量積累[11]。微擬球藻在氮源不足時,細胞蛋白與氨基酸含量顯著下降,碳代謝流向碳水化合物或者脂質從而提高糖類或油脂積累量[12]。萊茵衣藻在低氮脅迫條件下,淀粉含量和油脂合成之間存在著競爭關系[13]。國內外對低氮脅迫下微藻的代謝研究鮮見涉及蛋白核小球藻藻種,報道多集中于微藻生長和胞內生化組分方面,尚缺乏自養和葡萄糖兼養營養方式來源的藻細胞對低氮脅迫響應的系統比較,而且對胞外聚合物質 (extracellular polymeric substances,EPS) 研究極少。葡萄糖不僅是細胞的主要能量來源,還可以作為信號觸發特定的信號轉導通路控制細胞代謝狀態[14],葡萄糖兼養藻細胞對低氮脅迫處理的生化反應是否與自養藻細胞存在差異值得探究。微藻EPS主要為多糖、蛋白質等代謝產物,積累在細胞表面,通過穩定的膜結構來保護細胞,與藻細胞絮凝性能密切相關,并且在營養缺失時能為細胞提供碳與能量供應[15]。由此推測培養條件的改變可能通過影響EPS組成進而影響細胞絮凝收獲性能。由于微藻細胞尺寸小,當前普遍采用的離心沉降等物理收獲方法成本約占微藻生產總成本的三分之一,嚴重限制了其生產應用[16],因此研究低氮脅迫對藻細胞EPS組分及絮凝性能的影響具有重要現實意義。系統比較研究自養和兼養來源的蛋白核小球藻在生長速率、油脂含量、胞內外蛋白和淀粉等生化組分對低氮脅迫處理的響應差異,從EPS組分變化角度初步探究低氮脅迫處理對藻細胞絮凝性能的影響和相關機制,為蛋白核小球藻的人工培養技術優化提供科學依據。
1 材料和方法
1.1 供試材料
蛋白核小球藻藻種購自中國科學院武漢水生生物研究所,藻種編號為FACHB-1222。
1.2 主要儀器與試劑來源
電熱鼓風干燥箱101-3購自上海申光制造儀器公司;智能人工氣候箱safe PRX-600B購自寧波賽福實驗儀器有限公司;臺式冷凍恒溫振蕩器THZ-C-1購自蘇州培英實驗設備有限公司;電熱恒溫水浴鍋XMTD-8222購自上海精宏實驗設備有限公司;臺式高速離心機H1650購自湘儀儀器有限公司;超速冷凍離心機Centrifuge 5417R購自北京伯樂有限公司;顯微鏡BM1000購自江南永新;熒光光度計HitachiF-7000型購自日立高新技術公司。葡萄糖、NaNO3等生化試劑購自上海生工公司。
1.3 試驗方法
1.3.1 小球藻的培養和生長指標測定 以BG11培養基[17]為基礎培養基,挑取單藻落純化后接入BG11培養至對數期作為種子細胞,分別接入BG11自養培養基和添加10 g/L葡萄糖的BG11兼養培養基,接種初始濃度均為2 × 106cell/mL。待培養至穩定期,收獲藻細胞接種到低氮培養基,藻細胞接種后初始濃度均為1 × 107cell/mL。培養條件為25℃,光強2000 Lux,光周期中光暗比L∶D為12 h∶12 h,每天早晚各搖動一次。每組設置3個平行,自養和兼養來源藻細胞的培養周期均為5 d。BG11基本培養基氮素為18 mmol/L硝酸鈉 (1N),作為對照;以添加3種不同初始濃度 (3、6、9 mmol/L氮濃度,即,1/6N、1/3N、1/2N) 的硝酸鈉作為低氮脅迫處理培養基。
接種培養后每天取樣一次,取樣時間為光照10 h后,離心收集適量藻細胞并烘干至恒重,計算干重生物量。
1.3.2 藻細胞油脂提取和含量的測定 采用溶劑提取法和脂染色法對蛋白核小球藻油脂含量進行測定,并建立油脂含量與脂染色后在645 nm吸光度(A) 的線性回歸方程,采用脂染色法測定培養過程中的油脂。溶劑提取法操作參考Zhao等[18]的方法,脂染色法具體操作參照任潔等[19]的方法。油脂產率按照如下公式計算:
藻細胞油脂產率[mg/(L·d)] = 收獲藻細胞干重(mg/L) × 油脂含量/培養天數 (d)
1.3.3 小球藻胞內蛋白的提取和測定 在PBS緩沖液中冰浴條件下以超聲波破碎藻細胞后,用0.1 mol/L的NaOH振蕩提取2 h,8000 r/min離心15 min,收集上清,即為總蛋白。采用Bradford法[20]以牛血清白蛋白作標準測定蛋白濃度,換算出每克干重藻細胞總蛋白質含量。總蛋白含量減去藻細胞EPS中蛋白含量即為胞內蛋白含量。
總蛋白含量 (mg/g,藻細胞干重) = 蛋白濃度(mg/mL) × 提取液體積 (mL)/藻細胞干重 (g)
1.3.4 小球藻胞內淀粉的提取和測定 參照徐嬪等[21]的方法,以淀粉標準溶液測定繪制的標準曲線計算樣品中淀粉含量。藻細胞總淀粉含量減去胞外聚合物 (EPS) 中淀粉含量即為胞內淀粉含量。
1.3.5 小球藻胞外產物EPS的提取與分析 小球藻EPS的提取與分析參照Yang等[22]的方法。培養結束后,取藻細胞干重相當于0.1 g的微藻懸浮液,8 000 r/min離心10 min,棄上清,用5 mL去離子水洗滌沉淀,8 000 r/min離心5 min,微藻沉淀用5 mL去離子水懸浮并加熱至80℃,水浴30 min。隨后以8 000 r/min離心10 min,得上清液用0.45 μm醋酸纖維素膜過濾,濾液即EPS組分。按如1.3.4所述測定EPS組分中的胞外蛋白質含量和以淀粉含量表征的胞外多糖含量。
小球藻EPS三維熒光光譜 (3D-EEM) 的測定條件參照Lv等[23]的方法。使用150 W氙弧燈為激發光源,光電倍增管 (PMT) 電壓為700 V;設定激發狹縫寬度為10 nm,發射狹縫寬度為1 nm。光譜儀掃描范圍為激發波長/發射波長 (EX/EM) = 200~450 nm/250~550 nm,掃描速度為1200 nm/min。得到每個EX/EM所對應的熒光強度 (Intensity,單位為arbitary units,a.u.),對所有數據點采用Origin9.0軟件進行處理,形成等高線熒光光譜圖。
1.3.6 藻細胞絮凝率測定 參照Alam等[24]的方法并稍作修改。測定時搖動培養液以使藻細胞分散均勻,然后將10 mL細胞懸浮液轉移到試管中搖勻并測定藻細胞總濃度A (cell/mL),將培養液靜置12 h后,在距離液面頂部4 cm處取細胞懸液試樣,測量試樣藻細胞濃度B (cell/mL)。絮凝率按照如下公式計算:
絮凝率 (flocculating ability) = (A-B)/A × 100%
1.4 數據處理與統計分析
每個試驗處理設3次重復,各指標測定結果以“平均數 ± 標準誤”表示,應用SPSS22.0軟件對對照和各處理組數據進行方差分析,P< 0.05為差異顯著,P< 0.01為差異極顯著。采集數據利用Origin 9.0軟件進行分析作圖。
2 結果與分析
2.1 葡萄糖兼養培養對蛋白核小球藻生長和油脂積累的影響
表1顯示,自養與兼養的蛋白核小球藻分別在培養15 d和10 d時生長達到穩定期,此時兼養組的藻細胞干重生物量、油脂含量同比是自養組的1.62~2.82倍,油脂產率達到自養組的7.60倍。可見葡萄糖兼養培養使藻細胞生長周期縮短,產油效益大幅度提升。
2.2 低氮脅迫對不同來源蛋白核小球藻生長和油脂積累的影響
不同濃度低氮脅迫處理,自養和兼養來源的蛋白核小球藻生長和油脂積累如圖1所示。自養和兼養來源的蛋白核小球藻均在1/3N培養基中生長最快,4 d后進入穩定期,藻細胞干重分別達到2.56 g/L和4.62 g/L,分別為對照組 (1N) 的1.13倍和1.18倍,差異顯著。

表 1 自養與兼養培養對蛋白核小球藻生長和油脂積累的影響Table 1 The effect on growth and lipid accumulation of Chlorella pyrenoidosa of autotrophic and mixotrophic culture
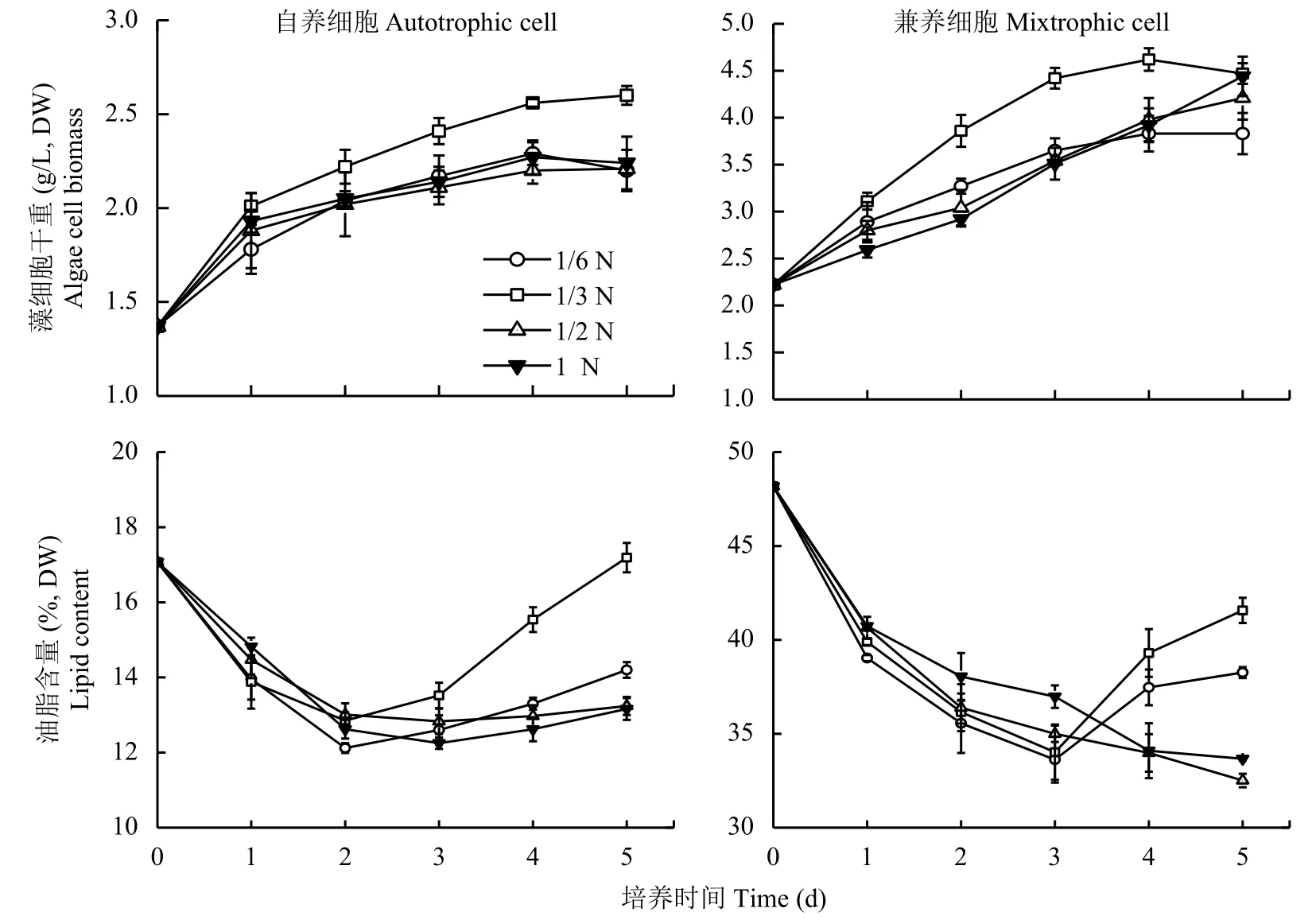
圖 1 不同濃度低氮脅迫下蛋白核小球藻干重和油脂含量Fig. 1 Dry biomass and lipid contents of Chlorella pyrenoidosa under low N stresses
隨著低氮脅迫處理時間的延長,自養和兼養來源的藻細胞油脂含量變化趨勢相似,1/2N和1N處理組藻細胞油脂含量下降后回升不明顯,1/6N和1/3N低氮脅迫的藻細胞油脂含量先下降后回升,具有明顯的拐點現象 (圖1),其中1/3N處理組油脂含量顯著高于其它組,可見1/3N低氮脅迫最能激發細胞的油脂合成。自養和兼養藻細胞油脂含量回升拐點分別出現在2 d和3 d,說明兼養來源的藻細胞對低氮脅迫響應較慢。培養至5 d時,1/3N處理的自養組藻細胞油脂含量為17.19%,與種子藻細胞初始值相當,但兼養組藻細胞油脂含量仍顯著低于其種子細胞初始值。
綜合藻細胞干重與油脂含量兩個指標計算油脂產率,自養與兼養組的蛋白核小球藻均是在1/3N低氮培養基中油脂產率最大,4 d油脂產率分別達到峰值 16.30 mg/(L·d) 和 129.56 mg/(L·d),同比 1N 對照組增幅分別為12.65%和35.82%,均顯著高于處理前藻細胞。可見,自養組藻細胞雖然對低氮脅迫的響應較敏感,但低氮脅迫對兼養組藻細胞油脂產率的提升效果更顯著。
2.3 低氮脅迫對不同來源蛋白核小球藻胞內蛋白和淀粉含量的影響
為了研究低氮脅迫處理對小球藻細胞的油脂合成與胞內其他生化組分的相關性,以1N組為對照,比較分析了1/3N低氮脅迫處理 (簡稱低氮組) 對自養和兼養來源的蛋白核小球藻胞內蛋白和淀粉含量的影響 (圖 2)。
自養和兼養來源的藻細胞胞內蛋白初始含量不同,兼養組藻細胞胞內初始蛋白含量 (152.07 mg/g)比自養組藻細胞高53.9%。轉接后第1天兼養組藻細胞胞內蛋白含量下降幅度均超過20.0%,高于自養組藻細胞的下降幅度 (8.0%以下)。自養組藻細胞胞內蛋白含量在4天內均逐日下降,只是低氮組每天降幅 (8.0%~10.0%) 高于對照組 (3.0%~6.0%),且最大降幅出現第3天,比對照組 (第2天) 晚。兼養組藻細胞對低氮脅迫處理的反應有所不同,1N對照組前3天胞內蛋白含量持續下降至谷值99.22 mg/g,第4天顯著回升至132.17 mg/g。可見在氮素供應充足(1N) 時,儲能相對豐富的兼養組藻細胞在轉接入新鮮培養基渡過適應性初期后,胞內蛋白即開始合成積累。相比而言,接入1/3N低氮培養基的兼養組藻細胞胞內蛋白含量呈持續下降趨勢。
自養組藻細胞胞內淀粉含量在轉接后先下降后上升,均于第3天達到谷值,與對照組相比,低氮組胞內淀粉含量降幅較大,第4天升幅更小。兼養來源的藻細胞胞內淀粉含量于轉接后第1天顯著下降,第2~3天維持在相對低值,之后明顯回升,4天時低氮組藻細胞胞內淀粉含量回升幅度也顯著低于對照組。
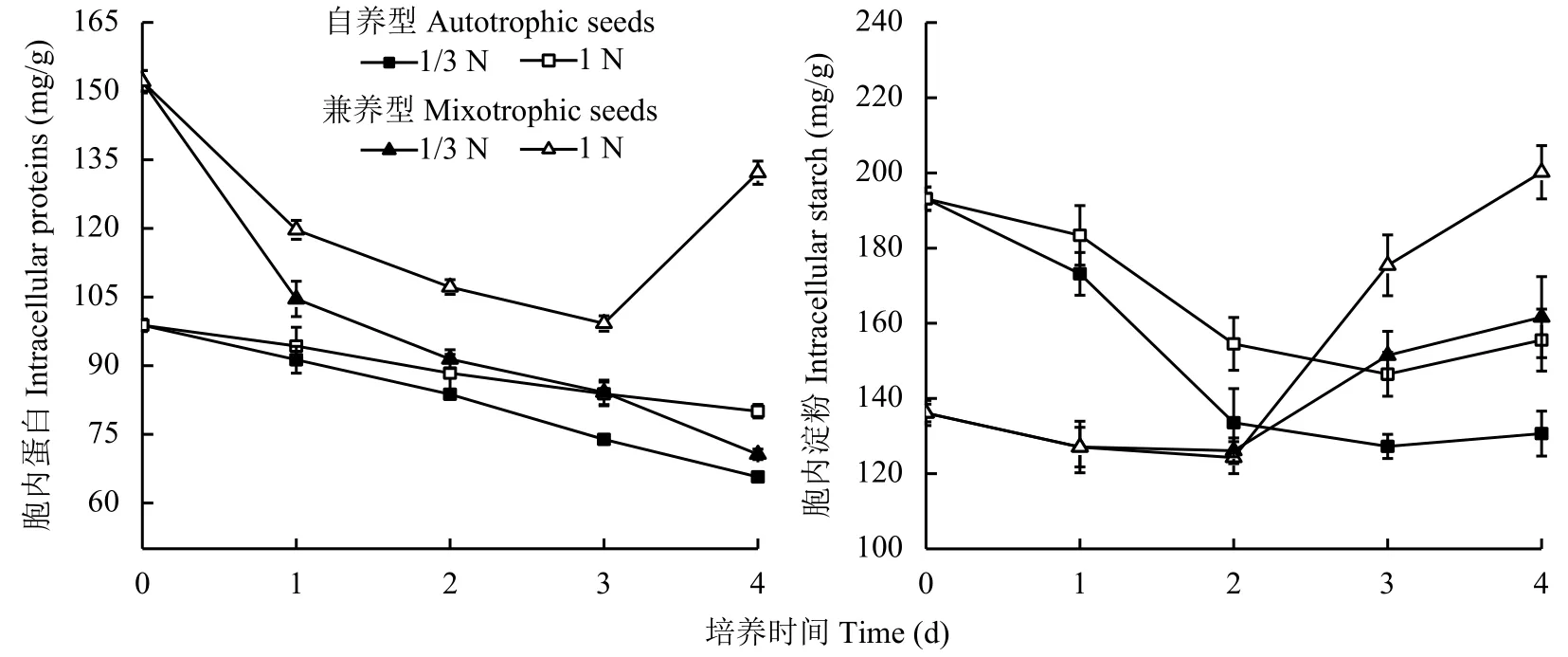
圖 2 培養基供氮水平1 N和1/3 N時蛋白核小球藻胞內蛋白和淀粉含量變化Fig. 2 Change incontents of intracellular proteins and starch of Chlorella pyrenoidosa grown in media with nitrogen supply levels of 1 N and 1/3 N
2.4 低氮脅迫對不同來源蛋白核小球藻胞外蛋白和多糖含量的影響
圖3表明,兼養組藻細胞EPS中蛋白含量顯著高于自養組,所有藻細胞轉接后EPS中蛋白含量均低于種子細胞的初始值。自養藻細胞對照組胞外蛋白含量持續下降,低氮組則前2天逐漸降至最低值2.69 mg/g,之后有所回升。兼養藻細胞EPS中蛋白含量均持續降低,低氮組同比對照組降幅更大,4天時低氮組藻細胞EPS蛋白含量比對照組低33.2%。
所有試驗組藻細胞EPS中多糖含量變化趨勢一致,均隨培養時間的延長而逐漸降低。兼養組藻細胞胞外多糖含量同比高于自養組,低氮組同比低于對照組。4天時兼養低氮組藻細胞EPS中多糖含量比對照組下降37.7%。
2.5 低氮脅迫對不同來源蛋白核小球藻EPS組分和絮凝性能的影響
2.5.1 對藻細胞EPS組分的影響 以1/3N低氮脅迫處理第4天的藻細胞為例,采用三維熒光掃描進一步比較了低氮脅迫處理對自養和兼養來源的藻細胞EPS組分的影響 (圖4)。自養與兼養組種子藻細胞EPS熒光掃描圖譜均在T、A和C區各存在一個主峰 (圖4A、B),分別代表不同類型的熒光基團。Chen等[25]對各峰的生化組分進行圖譜解析可知,T峰 (Ex/Em:280/352 nm) 為蛋白樣類色氨酸物質;A峰 (Ex/Em:230/373 nm) 是以富里酸為代表的類腐殖酸物質;C峰 (Ex/Em:330/373 nm) 是以胡敏酸為代表的類腐殖酸物質。T峰代表的蛋白樣類色氨酸物質中色氨酸屬于疏水性氨基酸,能促進藻細胞的絮凝,A和C峰代表的腐殖酸類物質具有親水性膠體性質,在高濃度時呈膠體溶液或分散體系[26],會抑制藻細胞絮凝。其中兼養組藻細胞EPS中T、A和C峰強度比顯著高于自養藻細胞。
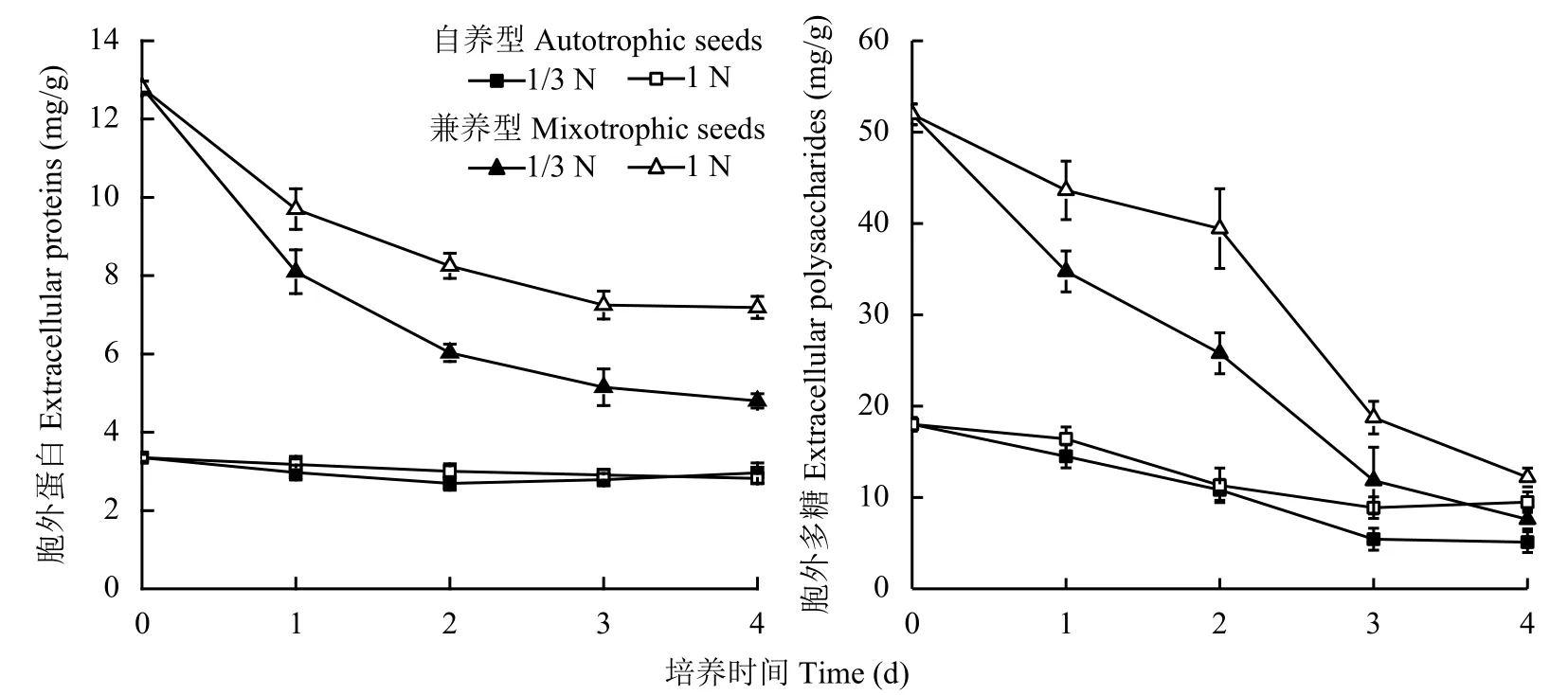
圖 3 培養基供氮水平1 N和1/3 N時蛋白核小球藻胞外蛋白和多糖含量變化Fig. 3 Change in contents of extracellular proteins and polysaccharides of Chlorella pyrenoidosa grown in media with nitrogen supply levels of 1N and 1/3N
與轉接前種子藻細胞 (圖4A) 相比,第4天自養對照組和低氮組藻細胞EPS熒光圖中A峰和C峰均消失,T峰強度明顯減弱 (圖4A1、A2),說明此時胞外類腐殖酸物質的含量極低,蛋白樣色氨酸物質含量也下降;但1/3N低氮組T峰強度比對照組增加40.3%,說明低氮脅迫處理能適當減緩藻細胞EPS中蛋白樣色氨酸含量的降幅。兼養來源的藻細胞轉接培養4 d后,EPS熒光掃描圖中T、A和C峰強度均顯著降低,其中1/3N低氮組T峰、C峰和A峰的強度分別是對照組的83.6%、74.8%和54.8% (圖4B、B1、B2),差異顯著,可見低氮脅迫組EPS中腐殖酸類物質含量降幅較大。
2.5.2 對藻細胞絮凝率的影響 藻細胞EPS組分含量的改變可能影響其絮凝率,測定自養和兼養來源種子藻細胞絮凝率和低氮脅迫處理4天后的絮凝率(表2)。兼養組藻細胞絮凝率顯著低于自養組。經1/3N低氮處理4天后,兼養和自養組藻細胞絮凝率為分別為78.46%和80.28%,均比對照組顯著提高。
3 討論
本結果表明,自養與葡萄糖兼養來源的蛋白核小球藻轉接入低氮培養基的第1天即開始生長分裂,3天后生長減緩 (圖1),說明低氮脅迫對藻細胞的促生長效應主要表現在培養初期。所有試驗組藻細胞轉接后初期油脂含量均顯著下降,隨著培養時間延長,1/6N和1/3N低氮脅迫處理組油脂含量出現先下降后回升的拐點現象 (圖1)。前人研究低氮脅迫提高藻細胞油脂含量的處理時間在5~12天之間,檢測間隔在1~3天之間[6,10,27],但一般藻細胞油脂含量隨培養時間而升高,未報道油脂含量升降拐點現象。原因可能在于不同藻種存儲油脂的代謝功用存在差異,油脂合成與分解代謝對培養環境變化的響應速率也不同。本文自養和兼養來源的藻細胞接入1/3N低氮培養基后油脂含量分別于第2天和3天出現回升拐點,說明自養組蛋白核小球藻種子細胞對低氮脅迫的響應速度比兼養組更為迅速。但葡萄糖兼養來源的藻細胞不僅初始油脂含量較高 (圖1),胞內外蛋白和淀粉 (多糖) 含量也較豐富 (圖2和圖3),1/3N低氮脅迫4天后油脂產率最高,同比是自養組藻細胞的7.95倍。說明種子細胞營養方式影響其對低氮脅迫處理的響應水平,而且藻細胞生物量增長與生化組分積累并不同步。Griffiths等[7]報道除螺旋藻外,小球藻、柵藻等10種微藻的生長速率和油脂含量均在1/10N低氮脅迫條件下最高。本研究中蛋白核小球藻的油脂含量在1/3N即6 mmol/L硝酸鈉低氮處理條件下效果最佳。
對蛋白核小球藻胞內外蛋白和淀粉 (多糖) 含量的監測數據表明,低氮組藻細胞胞內外蛋白、淀粉含量在培養初期均顯著下降,且降幅高于對照組 (圖2和圖3),這與Lee等[28]在萊茵衣藻C. reinhardtii上的研究結果相似。萊茵衣藻在氮限制條件下,氨基酸氧化酶的基因表達水平提高而淀粉的合成途徑將會被阻斷,原因在于藻細胞為了適應低氮環境,既加強了非必需蛋白的降解,也阻斷了淀粉合成對能量的消耗。但Fernandes等[29]研究卻發現,氮限制條件下凱式擬小球藻細胞內淀粉首先積累,待氮源耗盡之后,油脂含量開始逐步上升,說明淀粉是作為凱式擬小球藻碳與能量的初步儲存物質。本研究發現自養與兼養來源的藻細胞在低氮脅迫初期,油脂、淀粉和蛋白含量均迅速降低,與此相應的是生物量的大幅升高,待藻細胞生長逐步穩定的3天時,油脂和胞內淀粉作為儲備能源才開始逐步回升 (圖1和圖2)。而且兼養和自養種子藻細胞油脂含量 (圖1)和胞內淀粉含量高低變化同步,低氮脅迫處理后二者回升拐點出現時間也基本一致 (圖3),由此可推測淀粉和油脂均作為蛋白核小球藻的儲能物質,合成基本同步或淀粉合成后很快轉化為油脂儲存。至于蛋白質含量指標,除了兼養來源的藻細胞轉接入1N對照培養基3天后胞內蛋白含量明顯回升外,其他組藻細胞轉接后胞內外蛋白水平均維持在較低水平或持續下降,低氮脅迫組下降更為明顯 (圖2和圖3)。Geider等[30]報道三角褐指藻在氮脅迫條件下原初電子受體還原受阻而抑制了相關蛋白質的合成。Chen等[31]發現低氮脅迫后藻細胞蛋白質中部分氨基酸會發生分解代謝并能通過γ-氨基丁酸途徑、三羧酸循環再分配到脂質代謝途徑進行油脂合成。本研究蛋白核小球藻在1/3N低氮脅迫2天或3天后藻細胞油脂含量回升但胞內外蛋白含量持續下降,是否涉及蛋白降解氨基酸的轉化分配有待進一步探究。
微藻細胞的收獲成本在其生產成本中占比較大,藻細胞EPS組成與藻細胞絮凝收獲特性密切相關[32]。本研究所用蛋白核小球藻自養種子細胞EPS胞外蛋白和多糖含量較低 (圖3),二者比值也較低,而且含有一定比例的富里酸和胡敏酸類腐殖酸物質 (圖4A),這可能是其自絮凝率較低的原因。葡萄糖兼養的藻細胞EPS中對絮凝有促進作用的蛋白類色氨酸物質T峰強度比自養藻細胞增加34.3%,而不利于絮凝的腐殖酸類物質,A峰和C峰強度卻比自養組分別增加了141.8%和64.3% (圖4),這可能是葡萄糖兼養培養使藻細胞絮凝率下降的主要原因。與1N對照組相比,1/3N低氮組藻細胞EPS組分中蛋白樣類色氨酸和類腐殖酸物質均顯著下降,但T峰下降幅度低于A峰和C峰,從而顯著提高了藻細胞的絮凝率 (圖4和表2)。可見培養基組成的改變能影響微藻細胞EPS組分從而影響其絮凝收獲特性,在微藻人工培養技術中引入該項指標具有重要意義。本研究結果能為蛋白核小球藻人工培養技術的優化設計提供科學依據。
4 結論
蛋白核小球藻作為新食品資源開發的優良藻種,營養方式和低氮脅迫處理對其藻細胞生化組分積累轉化動態和收獲性能都具有顯著影響,而且胞內外不同組分的含量變化與其生長速率并不同步。1/3N低氮脅迫提高蛋白核小球藻的油脂產率和絮凝性能的效果最佳。
猜你喜歡
食品工業(2023年12期)2023-12-12 12:56:16
糧油食品科技(2022年6期)2022-11-25 12:18:34
糧油食品科技(2022年6期)2022-11-25 12:18:30
基層中醫藥(2020年8期)2020-11-16 00:55:20
環境衛生工程(2020年3期)2020-07-27 01:19:22
瘋狂英語·新讀寫(2020年3期)2020-06-06 09:06:14
塑料助劑(2019年3期)2019-07-24 08:51:22
中國糧油學報(2019年4期)2019-07-12 09:06:38
食品與機械(2018年11期)2019-01-03 06:15:34
中國糧油學報(2018年12期)2018-01-26 13:32:04
