秀水134為背景的染色體片段代換系的構建
2019-04-11 06:31:22孟卓玲曹玉潔單丹丹方春繆小菲費月新方柄杰王越趙文佳侯璐燕吳敏吳洪愷
中國稻米 2019年2期
孟卓玲 曹玉潔 單丹丹 方春 繆小菲 費月新方柄杰 王越 趙文佳 侯璐燕 吳敏 吳洪愷
(浙江農林大學 農業與食品科學學院,杭州311300;#共同第一作者;*通訊作者:hongkaiwu@163.com)
水稻是我國的主要糧食作物,世界上一半以上的人口以稻米為主食。水稻也是主要糧食作物中基因組較小的作物。隨著分子生物學和基因組學的發展,水稻作為模式生物,一方面,大規模創造突變體庫;另一方面,構建遺傳分析群體,開展了廣泛的分子生物學和基因組學研究。
性狀遺傳分析的基本方法是,一對相對性狀雜交構建分離群體,根據目標性狀在分離群體中的表現,推斷目標性狀的遺傳特征。隨著分子生物學和基因組學的發展,重要農藝性狀基因或QTL(Quantitative Trait Locus)的定位和克隆成為遺傳學研究的重要內容,遺傳分析群體同時又作為基因定位群體。最常用的群體有F2群體和回交F1群體,這類群體構建簡單,工作量小,時間短,但目標性狀檢測只能以單株(個體)進行,每種基因型僅由1個單株(個體)提供表型值,對于受環境影響較大的性狀,如產量,性狀值度量可靠性差。F2∶3群體,即用F2群體中1個個體衍生的F3群體(一般10~30個個體)的平均值代表1個F2個體的表型值,可以減小性狀值度量的誤差[1]。但與F2群體和回交F1群體一樣,個體中有雜合基因型,不能設置重復來估計或減小誤差。重組自交系群體(Recombination Inbred Lines,RIL)和加倍單倍體群體(Double Haploids,DH),這類群體的各個株系的基因型都是純合的,可以多年多點種植,通過設置重復來減小或估計誤差,提高了目標性狀鑒定的準確性。但這類群體的遺傳背景復雜,個體間遺傳差異大,對復雜的遺傳性狀來說,“遺傳噪音”大,基因定位的準確性低。
Paterson等[2-3]提出了構建染色體片段代換系(Chromosome segment substitution lines,CSSLs)用于基因精細定位和圖位克隆研究。CSSLs每個系與受體親本的遺傳差異僅限于某一染色體的代換片段,它們的表型差異只是由該代換片段的差異所引起,能有效地消除遺傳背景的干擾。CSSLs可以提高復雜農藝性狀基因定位的精確性,尤其是遺傳效應較小的數量性狀基因。在初步定位的基礎上,含有目標QTL的染色體片段代換系與受體親本雜交,建立次級分離群體,使QTL定位分析錨定在很窄的染色體片段上,進一步消除遺傳背景的干擾,從遺傳上和統計上提高了QTL定位和效應估計的準確性。日本水稻基因組研究計劃Yano研究組構建了以Nipponbare為遺傳背景的Kasalath全基因組染色體片段代換系,并將水稻抽穗期QTLHd-1、Hd-2、Hd-6和Hd-8分別分解為單個孟德爾因子,將Hd-3分解為2個單基因Hd-3a和Hd-3b,且精細定位了Hd-1、Hd-2、Hd-3a、Hd-3b、Hd-6和Hd-8等 6 個基因[4-5];在此基礎上,圖位克隆了Hd-1、Hd-3a和Hd-6[6-8]。日本九州大學Yoshimura研究組構建了分別以Asominori和IR24互為遺傳背景的兩套全基因組CSSLs[9-10]和一套以Taichung65為遺傳背景的非洲稻全基因組CSSLs[3]。Doi等[11]利用該CSSLs將第10染色體上抽穗期主效QTL分解為單基因Ehd1,并成功進行了圖位克隆。

圖1 秀水134和揚稻6號間多態性分子標記連鎖圖譜
以上這些研究成果和染色體片段代換系在有利基因發掘方面的優越性,極大地鼓舞了國內研究者開展構建染色體片段代換系的構建工作[12-13]。
1 材料與方法
1.1 材料
秀水134,浙江省推廣面積最大的晚粳稻品種,作為背景親本;揚稻6號,超級稻親本,為供體親本。
1.2 方法
1.2.1 多態性分子標記的篩選和連鎖圖譜的構建
根據McCouch構建的分子標記連鎖圖譜[14],選擇分子標記進行秀水134和揚稻6號間多態性分析;用Mapchart 2.2繪制多態性SSR標記連鎖圖。
1.2.2 染色體片段代換系構建
分3步進行:
第1步,親本秀水134和揚稻6號雜交并自交獲得F2群體;從F2群體中隨機選擇單株與秀水134回交,獲得BC1F1群體;BC1F1群體中隨機選單株與秀水134回交,獲得BC2F1群體;對BC2F1群體進行分子標記基因型鑒定,根據分子標記基因型,選擇代換片段盡量少、目的片段盡量長的單株,初步構建覆蓋全基因組的代換系(雜合型)。

圖2 以秀水134為輪回親本和揚稻6號構建染色體片段代換系(雜合型)
第2步,每個代換系繼續與秀水134回交,得到BC3F1群體,進行分子標記基因型鑒定,根據分子標記基因型,選擇代換片段盡量少、目的片段盡量長的單株,構建覆蓋全基因組的代換系(雜合型)。
第3步,BC3F1群體自交,在后代群體中進行分子標記基因型鑒定。選擇代換片段盡量少、目的片段盡量長的純合單株,組成覆蓋全基因組的代換系(純合型)。
1.2.3 代換片段長度的計算
按Young等[15]的方法計算代換片段的長度,不考慮2個相鄰分子標記間發生雙交換事件。當相鄰標記的基因型和供體親本的基因型相同時,認為這2個分子標記覆蓋的染色體區段為供體的代換片段;當相鄰標記基因型分別和供體親本、受體親本相同時,認為這2個標記之間的中點為該代換片段的邊界點,兩端邊界點之間的距離就是該代換片段的長度。
2 結果與分析
2.1 SSR分子標記連鎖圖譜的構建
根據McCouch等[14]構建的分子標記連鎖圖譜,從中選擇了629對引物,進行秀水134和揚稻6號間多態性分析,篩選到兩個親本間有多態的引物263對,親本間多態性比例為41.8%。選擇PCR擴增條帶分辨清晰,且均勻覆蓋水稻12條染色體的95對多態性分子標記,用Mapchart2.2繪制多態性SSR標記連鎖圖(圖1),全基因組總長度為1 499.6 cM。
2.2 染色體片段代換系的構建
分3步進行:
第1步,親本秀水134和揚稻6號雜交并自交獲得F2群體;從F2群體中隨機選擇97個單株與秀水134回交,獲得BC1F1群體,共97個株系;BC1F1群體每個株系種植10株,隨機選1株與秀水134回交,獲得BC2F1群體,共97個株系;BC2F1群體中每個株系隨機選3株進行分子標記基因型鑒定,根據分子標記基因型,選擇代換片段盡量少、目的片段盡量長的單株,篩選了53株(另24個單株作為備份),初步構建覆蓋全基因組的代換系(雜合型),見圖2。
第2步,每個代換系繼續與秀水134回交,得到BC3F1群體,共53個系,1 350株。對1 350株進行分子標記基因型鑒定,根據分子標記基因型,選擇代換片段盡量少、目的片段盡量長的單株,篩選了198株,構建覆蓋全基因組的代換系(雜合型),原理同第一步,圖略。
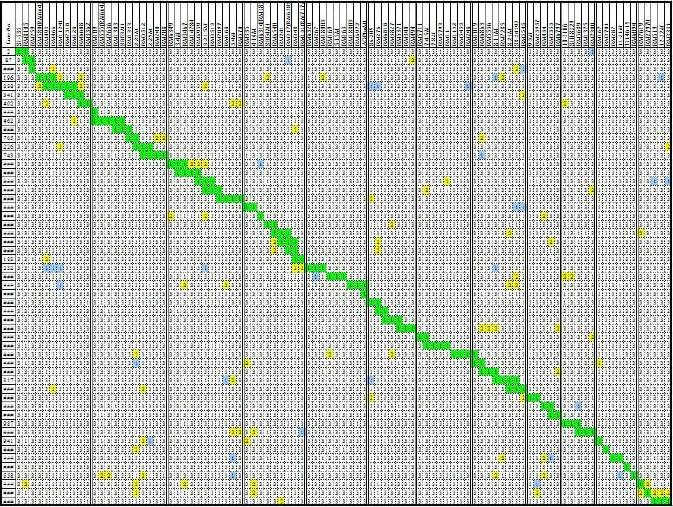
1代表揚稻6號基因型,2代表雜合型,3代表秀水134基因型。
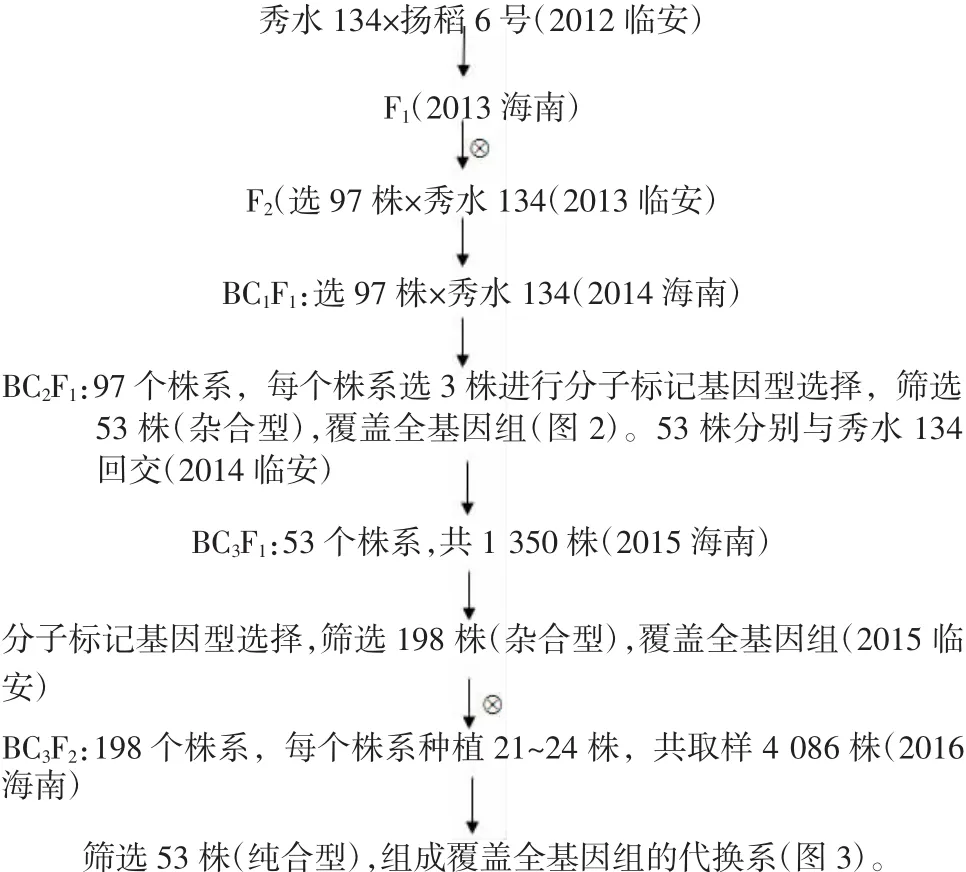
圖4 圖示染色體代換系構建流程
第3步,198株(雜合型)自交,得198個株系,每個株系種植21~24株,共取樣4 086株,進行分子標記基因型鑒定。選擇代換片段盡量少、目的片段盡量長的純合單株,組成覆蓋全基因組的代換系(純合型),見圖3。
以上3步的具體流程圖示為圖4。
2.3 染色體片段代換系的構建質量評價
對染色體片段代換系的53個株系的代換片段長度進行分析,代換片段最大的為7X5號株系,為182.15cM,占全基因組的 12.15%;代換片段最短的為7X29 號株系,為 7.4cM,占全基因組的 0.49%,見圖 5。該套染色體片段代換系代換片段總長度平均值75.30cM,占全基因組 5.00%。
12條染色體代換片段覆蓋總長度為1 995 cM,覆蓋水稻全基因組的1.33倍(表1)。其中,4號染色體的代換片段覆蓋總長度最長,為212.7cM,覆蓋4號染色體基因組的1.66倍;12號染色體代換片段覆蓋長度最短,為76.6 cM,覆蓋12號染色體基因組的0.77倍。除7號、10號、11號、12號染色體的代換片段沒有完全覆蓋各自的染色體基因組外,其余都覆蓋了整條染色體基因組。
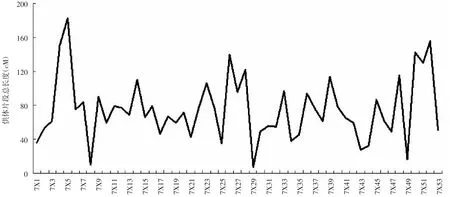
圖5 染色體片段代換系的代換片段長度的變異
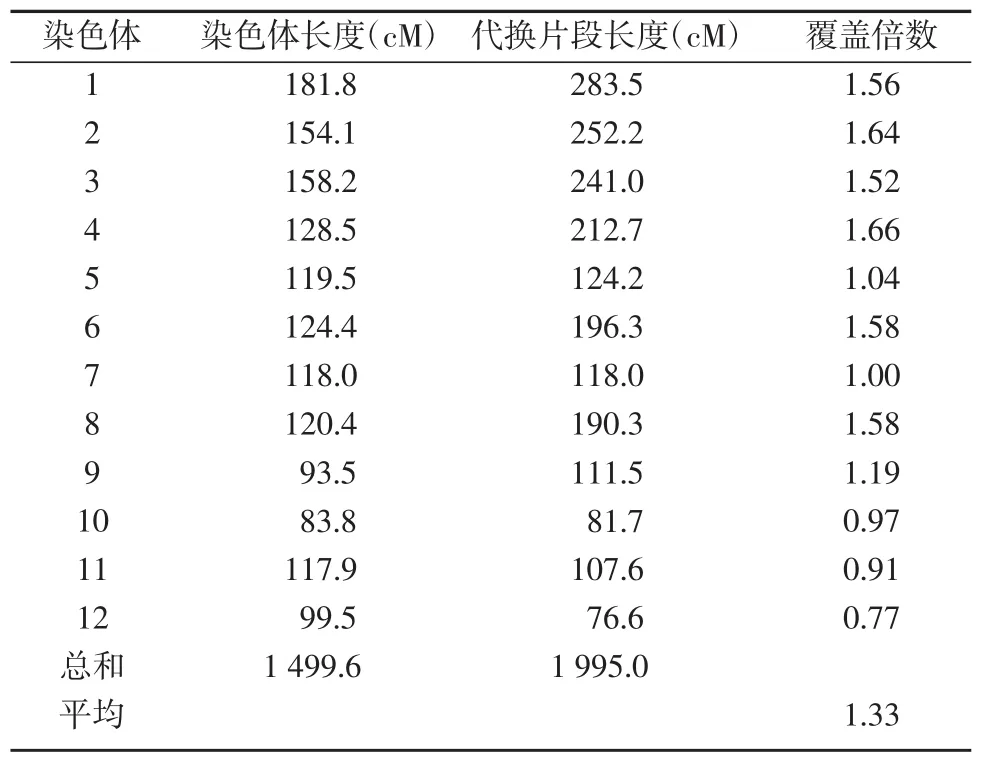
表1 染色體片段代換系各染色體目的代換片段長度
3 討論
3.1 染色體片段代換系構建的策略
染色體片段代換系作為發掘和鑒定有利基因的有力工具得到了廣泛的應用[16]。染色體片段代換系(chromosomal segment substitution lines,CSSL),又稱為染色體片段導入系(chromosomal segment introgression lines,CSIL)。基本的構建過程是:供體與受體雜交獲得F1;受體作為輪回親本,回交獲得BCnF1;自交,獲得目的位點基因型純合的個體,借助分子標記選擇,篩選來自供體的一個或多個代換片段的單株;所選單株的代換片段互相重疊,覆蓋全基因組,組成染色體片段代換系。可以采取兩種主要的策略:(1)BC1F1進行分子標記選擇,一開始就選定目的代換片段,組成完整的一套片段代換系,覆蓋整個基因組。不斷回交,在以后的世代中,一直選擇目的代換片段,同時進行背景選擇,直到獲得只有目的代換片段或帶少數幾個非目的代換片段的單株,所有這些單株的目的代換片段互相重疊,覆蓋全基因組,這樣就獲得了一套完整的染色體片段代換系。這樣的策略,獲得的代換片段長,但回交工作量大,耗時長。這是日本前期構建染色體片段代換系的策略。(2)在BC1F1群體中隨機選擇48株回交,得到48個株系,每個株系中隨機選擇1株回交,得到48個BC3F1株系,每個株系隨機選1株自交,得到48個BC3F2株系,每個株系選200~300株,進行分子標記基因型鑒定,篩選目的代換片段,下同方法1。這種方法,回交工作量小,耗時短,但分子標記鑒定工作量大。在染色體片段代換系構建過程中,可以把這兩種方法結合起來,做到取長補短。
3.2 染色體片段代換系的應用
以染色體片段代換系為材料可以提高對復雜農藝性狀基因定位的精確性,尤其適用于遺傳效應較小的數量性狀基因定位與克隆。在初步定位的基礎上,含有目標QTL的代換系與受體親本雜交,建立次級分離群體,使QTL定位分析錨定在很窄的染色體片段上,進一步消除遺傳背景的干擾,從遺傳上和統計上保證了QTL的精確定位。在明確控制重要性狀基因的基礎上,大力發掘和評價其等位性變異,可以開展設計育種[17]。這一過程中,利用已經鑒定出的各種重要育種性狀QTL的信息,包括QTL在染色體上的位置、遺傳效應、QTL之間的互作、QTL與背景親本和環境之間的互作等,模擬預測各種可能基因型的表現型,從中選擇符合特定育種目標的基因型,大幅度提高育種效率[18]。
本研究得到中國科學院遺傳與發育生物學研究所儲成才研究員的大力支持,謹致謝意!
