低溫脅迫對3種綠絨蒿生理生化指標的影響
2019-04-11 01:45:32侯禎丹冷秋思
西南農業學報 2019年3期
侯禎丹,屈 燕,黃 鴻,熊 健,冷秋思
(西南林業大學園林學院/園林植物與觀賞園藝省高校重點實驗室,云南 昆明 650224)
【研究意義】貝利葉綠絨蒿(Meconopsisbaileyi)、多刺綠絨蒿(Meconopsishorridula)和高莖綠絨蒿(Meconopsissuperba)屬罌粟科(Papaveraceae)綠絨蒿屬(Meconopsis),根據Gray Wilson (2012)最新的分類系統,分別來自Meconopsis,Grandes,Cumminsia3個屬,主要生長在高海拔地區的流石灘,高海拔地區空氣稀薄、氣溫低、溫差較大、日照輻射強烈,且大風襲擊使地表覆蓋大量的礫石層,表面土壤稀少,其幼苗在發育早期,必須忍受高山環境寒冷多變的氣候特征。因此,貝利葉綠絨蒿、多刺綠絨蒿和高莖綠絨蒿是研究高山植物在低溫脅迫下生理生化變化的理想材料。 【前人研究進展】綠絨蒿屬植物是云南著名的高山野生花卉,其花大、顏色豐富,具有巨大的遺傳育種潛力,同時還有清熱解毒、利尿、消炎、止痛等醫藥功效[1]。低溫及低溫脅迫時間的長短都會對植物體造成生理損傷,導致葉片表面呈水漬狀,使葉片容易受病菌的感染或造成植物體局部壞死,影響植物的正常生長[2]。任祝三[3]對總狀綠絨蒿、全緣葉綠絨蒿和粗壯綠絨蒿幼苗的生長氣候進行研究,結果表明幼苗生長受氣候條件影響較顯著。【本研究切入點】通過人工模擬低溫脅迫(4 ℃)不同時間下,對3種綠絨蒿幼苗葉片細胞的葉綠素含量、MDA含量、可溶性蛋白含量、游離脯氨酸含量及保護酶活性的變化進行分析,研究3種綠絨蒿在低溫處理不同時間的生理生化指標的變化規律。【擬解決的關鍵問題】研究低溫脅迫時間對不同綠絨蒿亞屬幼苗生理生化特性的影響,為綠絨蒿屬植物的引種栽培以及高山花卉植物抗低溫鑒定提供基礎理論依據。
1 材料與方法
1.1 試驗材料
以多刺綠絨蒿、貝利葉綠絨蒿、高莖綠絨蒿的半年生實生苗為材料,塑料花盆直徑9 cm、高9 cm,基質為蛭石+珍珠巖+泥炭土(1∶1∶7)的混合基質。置于溫度為20 ℃、濕度為75 %、光照設置為白天12 h、光照強度為120 μmol/m2·s夜間12 h、相對濕度保持在75 %~85 %的德國智能人工氣候培養箱中預培養6 d,在此期間根據生長情況加入適量的水分。
1.2 試驗方法
選取生長健康和長勢基本一致的幼苗6株,放置在德國智能型人工氣候箱內(Binder710,德國)進行低溫處理。光照強度為120 μmol/m2·s,光照時間8:00-20:00時,相對濕度保持在75 %~80 %,以20 ℃進行第0小時(對照)測定,設置人工氣候箱以4 ℃/h降溫,降至(4.0±0.5)℃保持該溫度為低溫處理溫度,在第24、48、72小時以及恢復20 ℃ 48 h (低溫解除,記為re48 h)進行各項生理指標測定。
1.3 葉片采集
葉片樣品采集時間為8:30 am,以綠絨蒿幼苗葉片的全展葉為材料,將蒸餾水洗凈的葉片磨碎進行不同生理指標測定,每個指標做6個重復。
1.4 生理指標的測定
葉綠素含量的測定用分光光度法;丙二醛(MDA)測定采用硫代巴比妥酸法;超氧化物歧化酶(SOD)活性的測定采用氮藍四唑(NBT)光化還原法;游離脯氨酸的測定采用磺基水楊酸法;可溶性蛋白含量采用考馬斯亮藍法測定。
2 結果與分析
2.1 葉綠素含量變化
由圖1可知,3種綠絨蒿幼苗葉片的葉綠素含量均呈現先降后升的趨勢。貝利葉綠絨蒿和高莖綠絨蒿在24 h下降幅度最大,分別為43.64 %和29.57 %,貝利葉綠絨和高莖綠絨蒿在低溫處理24 h后,隨著低溫處理時間的延長葉片的葉綠素含量逐漸呈上升趨勢;多刺綠絨蒿葉綠素含量則在48 h最低,下降幅度為23.07 %。恢復48 h后,貝利葉綠絨蒿和多刺綠絨蒿的葉綠素含量下降幅度為處理前的15.39 %和12.04 %,高莖綠絨蒿葉綠素含量上升幅度為處理前的6.71 %。
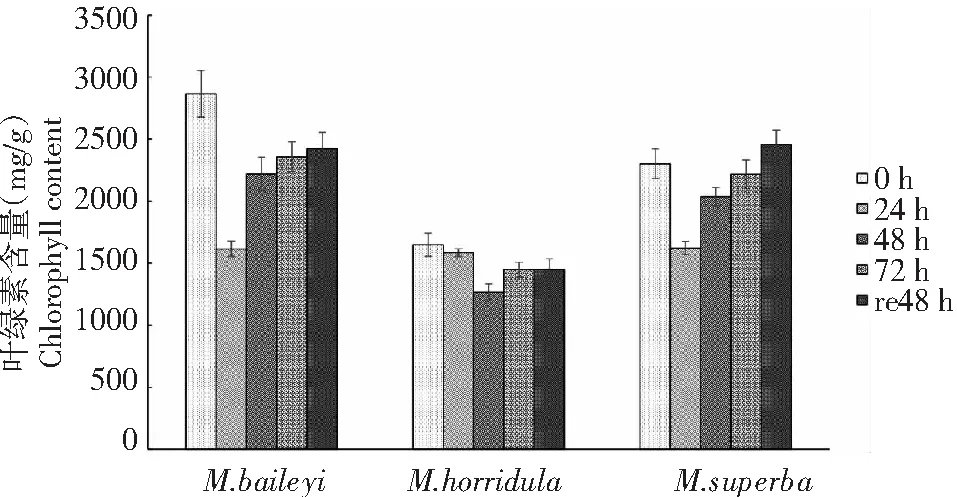
圖1 低溫脅迫下3種綠絨蒿幼苗葉綠素含量變化Fig.1 Changes of chlorophyll contents of three Meconopsis seedlings under low temperature stress
2.2 MDA含量變化
由圖2可知,3種綠絨蒿幼苗葉片MDA含量均呈先升后降趨勢。低溫處理24 h貝利葉綠絨蒿和高莖綠絨蒿上升較快,上升幅度分別為處理前的145.2 %和154.6 %,在低溫脅迫時間24 h后,這2種綠絨蒿的MDA含量均呈下降趨勢,恢復48 h后MDA的含量比處理之前高,上升幅度分別5.82 %和13.5 %;多刺綠絨蒿幼苗葉片在低溫脅迫時間為72 h時MDA含量最高,上升幅度為處理之前的67.85 %。
2.3 可溶性蛋白含量變化
由圖3可知,3種綠絨蒿的蛋白含量均呈先升后降趨勢,貝利葉綠絨蒿和多刺綠絨蒿的葉片蛋白含量在低溫處理24 h含量最高,上升幅度分別為53.82 %和103.27 %,在恢復48 h后貝利葉綠絨蒿和高莖綠絨蒿葉片可溶性蛋白含量上升幅度為處理前的11.92 % 和47.57 %;高莖綠絨蒿幼苗葉片可溶性蛋白含量處理72 h時最高,上升幅度為處理前的108.24 %,在恢復48 h后可溶性蛋白含量為處理前的47.57 %。

圖2 低溫脅迫下3種綠絨蒿幼苗MDA含量變化Fig.2 Changes of MDA contents of three Meconopsis seedlings under low temperature stress
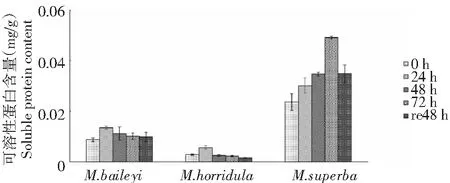
圖3 低溫脅迫下3種綠絨蒿幼苗可溶性蛋白含量變化Fig.3 Changes of soluble protein contents of three Meconopsis seedlings under low temperature stress
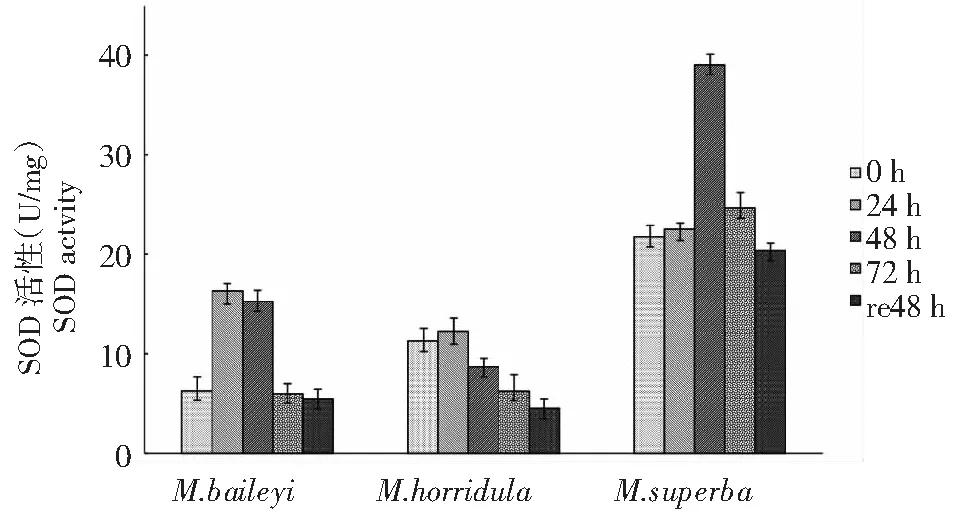
圖4 低溫脅迫下3種綠絨蒿幼苗SOD活性變化Fig.4 Changes of SOD activity of three Meconopsis seedlings under low temperature stress
2.4 SOD活性變化
由圖4可知,3種綠絨蒿幼苗葉片SOD活性均呈現先升后降的趨勢。高莖綠絨蒿幼苗葉片的SOD活性比較高,貝利葉綠絨蒿和多刺綠絨蒿的SOD活性較低;在低溫處理24 h時,貝利葉綠絨蒿和多刺綠絨蒿SOD活性最強,上升幅度分別為處理前的 159.16 % 和9.18 %,在低溫處理48 h后貝利葉綠絨蒿SOD活性迅速下降,貝利葉綠絨蒿和多刺綠絨蒿恢復48 h后SOD的活性均比處理前低,下降幅度為13.82 %和59.88 %;而在低溫處理48 h時,高莖綠絨蒿幼苗葉片SOD活性最強,上升幅度為處理前的79.78 %,之后隨低溫脅迫時間的延長呈下降趨勢,在低溫脅迫恢復48 h時,下降幅度為低溫處理前的6.15 %。
2.5 游離脯氨酸含量變化
由圖5可知,3種綠絨蒿幼苗葉片游離脯氨酸含量均呈現先升后降的趨勢,在低溫脅迫24 h貝利葉綠絨蒿和高莖綠絨蒿幼苗葉片游離脯氨酸含量上升幅度較快,上升幅度是處理前的118.75 %和445.13 %,到48 h后游離脯氨酸的含量呈下降趨勢,恢復48 h后游離脯氨酸的含量比低溫處理前低,下降幅度分別為4.17 %和41.59 %;而多刺綠絨蒿幼苗在低溫處理48 h前呈現上升趨勢,低溫處理48 h時上升幅度為140 %,之后葉片中游離脯氨酸含量開始下降,恢復48 h后游離脯氨酸含量上升幅度為處理前27.5 %。
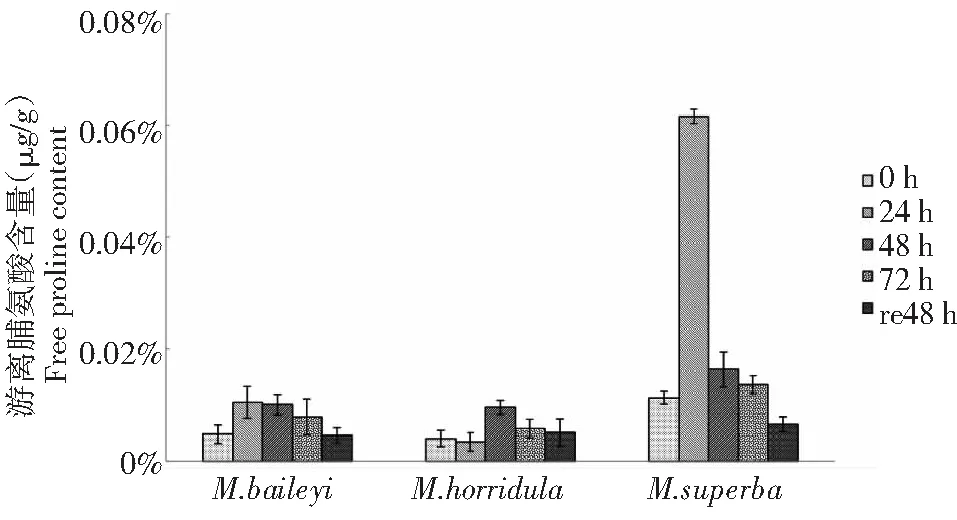
圖5 低溫脅迫下3種綠絨蒿幼苗游離脯氨酸含量變化Fig.5 Changes of free proline contents of three Meconopsis seedlings under low temperature stress
3 討 論
3.1 低溫脅迫時間與葉片葉綠素含量
葉綠體是植物進行光合作用的主要場所,逆境脅迫下部分光合機制遭到破壞,植物體內的葉綠素含量為適應逆境環境而產生生理變化[4]。光合作用是植物能量轉化的來源,植物對低溫最敏感的器官是光合器官,低溫又影響葉綠素的合成,更造成葉綠素降解加強,導致植物幼苗的光合減弱。張雪[5]研究發現,低溫下葉片內葉綠素含量略有增加,可能是由于葉綠素形成的最低溫度是2~4 ℃;張學財[6]等的研究發現在4 ℃處理時,3種劍麻品種葉綠素含量先降后升,這也可能是機體對低溫環境逐步適應的表現。本研究中3種綠絨蒿幼苗葉片在低溫脅迫下葉綠素含量均呈先降后升的趨勢,貝利葉綠絨蒿和高莖綠絨蒿幼苗葉片處理24 h葉綠素含量先大幅度下降再逐漸上升,多刺綠絨蒿處理48 h緩慢降到最低隨后上升,說明在低溫處理初期綠絨蒿幼苗葉片無法進行光合作用,導致幼苗無法合成葉綠素并開始降解,故葉綠素含量降低;隨后葉綠素含量逐漸上升,說明在持續4 ℃的低溫下,低溫沒有使葉綠素合成受到破壞,而是限制了葉綠素的合成。貝利葉綠絨蒿和高莖綠絨蒿幼苗葉片葉綠素含量處理24 h時最低,隨后葉綠素含量遞增,而多刺綠絨蒿處理48 h時葉綠素含量最低,而后逐漸升高,說明貝利葉綠絨蒿和高莖綠絨蒿在低溫處理下能更快速地對低溫逆境脅迫做出反應。
3.2 低溫脅迫時間與MDA含量
植物器官在低溫逆境條件下,會發生膜脂過氧化作用產生MDA。植物細胞質膜損傷的程度由MDA含量的變化反應,MDA在一定程度上也反映了植物抗逆性的強弱,葉片中MDA積累越少,膜質過氧化程度小,表明組織的抗逆性越強。田丹青[7]的研究發現,低溫處理12 h 后3個紅掌品種葉片的 MDA 含量均呈降低的趨勢,MDA含量隨著低溫處理時間的延長而增加,這與相對電導率的測定結果以及冷害癥狀表現一致。張玲麗[8]的研究發現14個百合品種在低溫脅迫1~3 d后所有百合品種MDA含量都有不同程度的升高,低溫脅迫3 d后,各品種的MDA含量呈極顯著差異。本研究低溫脅迫3種綠絨蒿的葉片MDA含量均呈現先升后降的趨勢,貝利葉綠絨蒿和高莖綠絨蒿幼苗在低溫脅迫24 h MDA含量呈先大幅上升再緩慢下降的變化趨勢,表明細胞膜系統受到損傷,保護酶清除速率低于活性氧產生速率,積累大量活性氧,導致膜質過氧化作用更加劇烈,MDA含量快速增加,嚴重阻礙了保護酶活性并降低抗氧化物的含量[9],迫使細胞膜系統損傷更加嚴重[10];多刺綠絨蒿幼苗葉片MDA含量隨低溫脅迫時間的延長呈上升趨勢且增幅小,說明其在低溫逆境條件下,膜質過氧化程度小,表明多刺綠絨蒿幼苗葉片組織的抗逆性強。貝利葉綠絨蒿和高莖綠絨蒿幼苗葉片處理24 h時MDA含量最高,隨后呈遞減趨勢,多刺綠絨蒿處理72 h 時MDA含量最高,且多刺綠絨蒿MDA含量增幅小。
3.3 低溫脅迫時間與可溶性蛋白含量
在逆境環境下植物通過可溶性物質的積累來調節滲透,可溶性蛋白含量的變化反應了植物合成和代謝能力,低溫條件下,可溶性蛋白含量的增加,有助于加強細胞的保水能力,從而提高植物體的抗寒性。抗寒性強的品種可溶性蛋白含量高,在低溫逆境環境中可溶性蛋白含量呈上升趨勢[11]。沙偉[12]等的研究發現在低溫脅迫下扎龍蘆葦葉片內可溶性蛋白的含量明顯下降,但仍高于對照組。在低溫脅迫下抗寒性強的葡萄品種可溶性蛋白含量高且增加幅度大[13];在低溫脅迫下隨溫度的降低蕓香可溶性蛋白含量增加[14];抗寒性強的葡萄品種可溶性蛋白形成早且含量高[15]。楊慧菊[16]等研究發現低溫脅迫下馬鈴薯幼苗的可溶性蛋白含量不斷上升。本研究貝利葉綠絨蒿和高莖綠絨蒿幼苗葉片在恢復48 h后可溶性蛋白含量下降,但仍高于處理前含量,細胞的持水力由可溶性蛋白的親水膠體性決定,可溶性蛋白含量的增加可以約束更多的水分,減少原生質因結冰而傷害致死的機率。貝利葉綠絨蒿和多刺綠絨蒿幼苗處理24 h可溶性蛋白含量最低,隨低溫脅迫處理時間延長可溶性蛋白呈下降趨勢,說明抗寒能力弱,而高莖綠絨蒿幼苗葉片內可溶性蛋白含量隨著低溫處理時間的延長明顯上升,處理72 h可溶性蛋白含量最高,說明低溫處理后高莖綠絨蒿幼苗葉片內可溶性蛋白可能是從膜上或其它的結合形式中降解釋放的,也可能是有新的蛋白質合成。
3.4 低溫脅迫時間與SOD活性
在植物體內多種酶維持活性氧在動態平衡的低水平,是植物對低溫逆境脅迫的初始反應。超氧化物陰離子自由基產生的H2O2和O2可以被SOD清除,從而防止活性氧過多積累對植物體造成傷害來維持活性氧的動態平衡。張尚雄等[17]研究表明3個披堿草屬牧草隨低溫脅迫的增強葉片SOD活性急劇增加,表明3個披堿草屬牧草受到寒冷脅迫時產生了氧自由基以抵御寒冷。本研究低溫脅迫下3種綠絨蒿SOD活性均呈現先升后降的趨勢,持續低溫脅迫誘導激發了酶的活性使酶活性增強,從而提高植物體細胞抗寒性,由于環境溫度過低導致酶的活性下降,從而使植物體細胞結構遭到破壞,活性氧被保護酶系統清除的能力下降,活性氧大量積累致使膜系統結構和功能受到破壞。貝利葉綠絨蒿和多刺綠絨蒿幼苗葉片SOD活性處理24 h活性最強,而高莖綠絨蒿處理72 h活性最強,增幅最大。
3.5 低溫脅迫時間與游離脯氨酸含量
游離脯氨酸是滲透脅迫下產生的滲透調節物質,能保護植物體細胞內生物大分子、穩定細胞蛋白質結構及保持氮含量的作用[18]。王靜[19]等研究認為,脯氨酸含量與植物抗寒性關系密切,低溫脅迫程度增加會使游離脯氨酸含量增加。鄧雪珂等[20]研究表明,紫花苜蓿隨低溫處理時間的延長游離脯氨酸含量大幅增加,在低溫脅迫12 h后游離脯氨酸含量增加十分顯著。羅新義等[21]發現,隨低溫處理時間的延長肇東苜蓿葉片中游離脯氨酸含量逐漸增加。本研究低溫脅迫下3種綠絨蒿幼苗葉片游離脯氨酸含量均呈現先升后降的趨勢,貝利葉綠絨蒿和高莖綠絨蒿在處理24 h時游離脯氨酸的積累達到峰值隨后下降,有可能是超過滲透調節能力范圍造成的生理損傷反應;多刺綠絨蒿隨低溫處理時間的延長呈上升趨勢,在處理48 h時達到峰值,游離脯氨酸的積累可能對植物體內滲透調節起到了作用。高莖綠絨蒿在處理24 h時游離脯氨酸含量較其它兩種綠絨蒿增幅大。
4 結 論
通過比較3種綠絨蒿在低溫脅迫下,其生理生化指標對低溫的響應規律,發現3種綠絨蒿均具有一定的抗寒性,而高莖綠絨蒿較之其它兩種的抗寒能力更強。研究結果可為今后綠絨蒿屬植物雜交育種、抗寒基因篩選等工作提供參考。
