不同林齡第2代杉木林枝葉凋落前的養(yǎng)分轉(zhuǎn)移特征
2019-04-17 13:13:18呂中誠康文星黃志宏趙仲輝鄧湘雯
生態(tài)學(xué)報(bào) 2019年5期
呂中誠, 康文星, 黃志宏,趙仲輝,鄧湘雯
1 中南林業(yè)科技大學(xué),長沙 410004 2 南方林業(yè)生態(tài)應(yīng)用技術(shù)國家工程實(shí)驗(yàn)室,長沙 410004 3 國家野外科學(xué)觀測研究站,會(huì)同 418307 4 湖南警察學(xué)院,長沙 410138
植物組織衰老時(shí)養(yǎng)分向活體組織轉(zhuǎn)移,這種養(yǎng)分轉(zhuǎn)移屬于植物養(yǎng)分內(nèi)循環(huán)。這種養(yǎng)分轉(zhuǎn)移使得養(yǎng)分在植物體內(nèi)的存留時(shí)間延長,被植物再吸收、再分配和再利用[1- 3]。目前,許多學(xué)者在不同生活型植物葉片凋落前養(yǎng)分轉(zhuǎn)移率的差異[4- 7],不同林齡植物葉片凋落前養(yǎng)分轉(zhuǎn)移的變化規(guī)律[8- 13],不同氣候和氣候變化對植物葉片凋落前養(yǎng)分轉(zhuǎn)移的影響[14],土壤肥力、施肥、溫室條件下和養(yǎng)分有效性對養(yǎng)分轉(zhuǎn)移的影響[15-16]等方面進(jìn)行了研究。也有學(xué)者探討了植物葉凋落前養(yǎng)分轉(zhuǎn)移的機(jī)理[17-18]。這些研究成果具有重要的實(shí)踐和理論意義。但是大部分研究主要集中在林分某一時(shí)間段的植物葉凋落前養(yǎng)分轉(zhuǎn)移,雖然也有研究不同林齡植物葉凋落前的養(yǎng)分轉(zhuǎn)移動(dòng)態(tài)[8- 13],卻都采用“時(shí)空轉(zhuǎn)換”法。由于立地條件異質(zhì)性,研究結(jié)果可能偏移實(shí)際,而且由于時(shí)空的異質(zhì)性,也不能真正體現(xiàn)某一林分從幼林到成熟林不同林齡的植物衰老組織凋落前的養(yǎng)分轉(zhuǎn)移規(guī)律。一個(gè)林分從幼林到成熟期不同年齡時(shí)植物衰老組織凋落前的養(yǎng)分轉(zhuǎn)移規(guī)律怎樣?是森林養(yǎng)分循環(huán)和養(yǎng)分利用深入研究中亟需解決的問題。
杉木(Cunninghamialanceolata(lamb) Hook)具有易繁殖、生長快、產(chǎn)量高、材質(zhì)好等特點(diǎn),是我國特有的重要速生用材樹種。據(jù)第八次全國森林資源清查結(jié)果,杉木栽植面積1.069×107hm2[19],在我國人工林業(yè)生產(chǎn)建設(shè)中具有舉足輕重的地位。本研究利用會(huì)同杉木林25年的定位測定的基礎(chǔ)數(shù)據(jù),從養(yǎng)分角度來探討杉木人工林經(jīng)營問題。目的是通過枝、葉凋落前的養(yǎng)分轉(zhuǎn)移研究,探討杉木人工林養(yǎng)分內(nèi)循環(huán)隨林齡的變化規(guī)律,比較不同年齡杉木對養(yǎng)分再吸收利用的能力,為杉木人工林的經(jīng)營管理提供理論依據(jù)。
1 材料與方法
1.1 研究區(qū)概況
本研究在我國杉木中心產(chǎn)區(qū)之一湖南會(huì)同深沖林場(中國國家野外科學(xué)觀測研究站—湖南會(huì)同杉木林生態(tài)系統(tǒng)研究站的科研基地)第3小集水區(qū)進(jìn)行。地理位置 26°50′N,109°45′E,海拔270—350 m,坡向西北,坡度15—25°。研究區(qū)屬于中亞熱帶季風(fēng)濕潤氣候區(qū),年平均氣溫16.9℃,年降雨量1100—1400 mm,年均相對濕度80%以上。土壤母巖為震旦紀(jì)板溪系灰綠色板巖,風(fēng)化程度較深,土層厚度80 cm以上,土壤為中有機(jī)質(zhì)厚層黃壤。研究林分為1988年春在1987年冬皆伐第1代杉木林的采伐跡地上,采用煉山、全墾挖穴造林方式,營造的第2代杉木林。造林密度2200 株/hm2,杉木栽上后前3年的春秋季節(jié)各撫育1次,以后任其自然生長。不同林齡時(shí)林分結(jié)構(gòu)特征見表1。
1.2 樣品采集
中南林業(yè)大學(xué)會(huì)同生態(tài)研究站為了研究杉木林生態(tài)系統(tǒng)的結(jié)構(gòu)功能,在第3小集水區(qū)第2代杉木林栽下后,在林內(nèi)設(shè)立了4塊觀測樣地(每塊樣地面積667 m2)進(jìn)行各項(xiàng)觀測試驗(yàn)(小集水區(qū)形狀為兩山坡夾一山槽,面積1.98 hm2,在每山坡的中上部和中下部各設(shè)1塊觀測樣地,共計(jì)4塊觀測樣地,位置見圖1)。林分7、11、16、20年生和25年生時(shí),在4塊觀測樣地測定林分生物量。每次測定生物量的同時(shí)采集養(yǎng)分待測樣品。為了保證樣品具有代表性,在每一次采集樣品的伐倒的標(biāo)準(zhǔn)木(依據(jù)樣地的平均樹高、平均胸徑定的標(biāo)準(zhǔn)木)上把它分成樹干基部、中下、中、中上和上部5個(gè)區(qū)段,每一區(qū)段收集相同重量的多生枝、一年生枝、當(dāng)年生枝混合一起組成活枝樣品,活樹葉采用樹枝相同的方法。每一樣地在林下安置兩個(gè)面積為18—20 m2的塑料薄膜裝置承接枝、葉凋落物。每月中旬和月底收集枝、葉調(diào)落物及其待測樣品(待測樣品盡可能收集剛掉落的枝葉)。
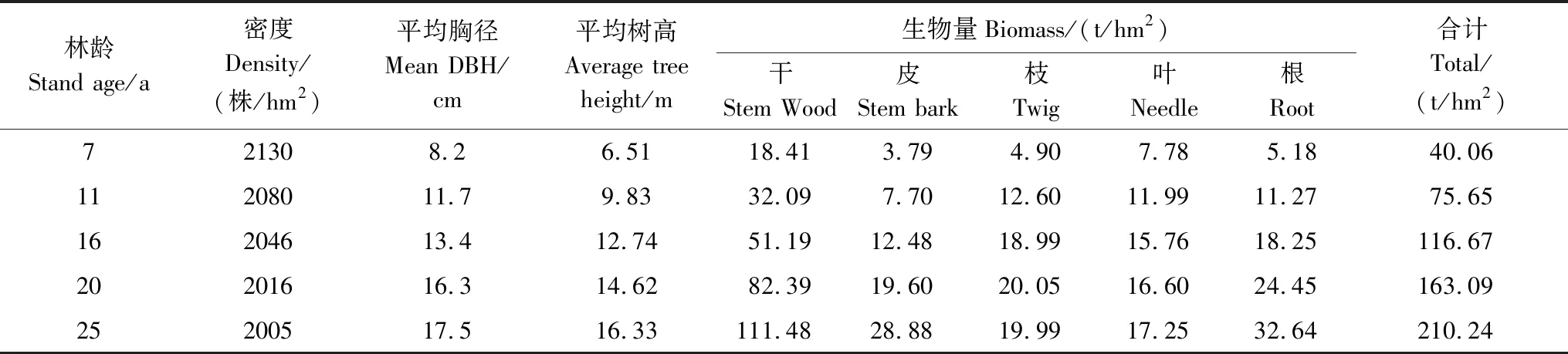
表1 不同林齡杉木林的林分特征
圖1 觀測樣地位置 Fig.1 The observation site location
1.3 不同林齡段枝葉枯死量估算
每月中旬和月底收集塑料薄膜裝置承接枝、葉凋落物,將凋落枝、葉樣品置于烘箱在80℃溫度下將樣品烘干至恒重,求算含水率,通過換算將凋落枝、葉鮮量換算成干質(zhì)量。根據(jù)凋落物收集的大薄膜裝置面積和林地面積換算,求算出樣地林分每月的枝葉凋落量,再將1年各月累計(jì)估算1年的林分枝葉凋落量。
1.4 養(yǎng)分測定
將待測杉木活的和死的枝、葉干物質(zhì)樣品磨碎,過60目篩后,按《森林土壤定位研究方法》[20]配置待測溶液。植物中的全N用半微量凱氏法;全P用分光光度計(jì)測定;全K、全Ca、全Mg均用原子吸收分光光度計(jì)測定。每個(gè)樣品重復(fù)測定3 次,用3 次重復(fù)測定的平均值作為計(jì)量標(biāo)準(zhǔn)。本研究中活的枝、葉的養(yǎng)分濃度是在7、11、16、20年生和25年生時(shí),測定林分生物現(xiàn)存量采集的活枝葉樣品測定的,凋落枝葉的養(yǎng)分也是在測定林分生物量的同時(shí)收集的凋落枝葉樣品測定的。而且,由于植物組織養(yǎng)分濃度是逐漸變化的,如11年生測定的養(yǎng)分濃度為K,16年生時(shí)測定的為W,若W-K=1,將1除以11年至16年生這林齡段的年數(shù)(5年),那么12年生時(shí)養(yǎng)分濃度為K+0.2,13年生時(shí)為K+0.4,依次類推16年生為K+1=W。再將每年增加的值相加除以這林齡段的年數(shù)后,再加上11年生測定的養(yǎng)分濃度值,作為11年至16年生這林齡段的年均養(yǎng)分濃度。
1.5 不同林齡段枝、葉落凋落前養(yǎng)分轉(zhuǎn)移量和轉(zhuǎn)移率估算
采用枝葉凋落前后的養(yǎng)分濃度之差及單位面積枝葉凋落量來計(jì)算養(yǎng)分轉(zhuǎn)移量。若某林齡段測定的杉木活枝、葉養(yǎng)分濃度為L1i,此林齡段測定凋落枝、葉的養(yǎng)分濃度為L2i。某林齡段的單位面積枝、葉凋落量為Wi,那么這林齡段杉木枝、葉在凋落前養(yǎng)分轉(zhuǎn)移量用公式(1)計(jì)算。養(yǎng)分轉(zhuǎn)移率采用周麗麗的研究方法[13],用公式(2)計(jì)算:
M=Wi×(L1i-L2i)
(1)
R=(L1i-L2i)/L1i×100%
(2)
式中,M表示林分某林齡段杉木枝、葉凋落前養(yǎng)分轉(zhuǎn)移量(kg/hm2);R為養(yǎng)分轉(zhuǎn)移率(%);i=1,2(枝、葉)。
1.6 數(shù)據(jù)分析處理
所有數(shù)據(jù)經(jīng)Excel軟件處理,用SPSS 20.0軟件進(jìn)行統(tǒng)計(jì)分析。釆用最小顯著差異法(LSD)對同林齡時(shí)活葉與死葉、活枝與死枝的養(yǎng)分濃度,各元素轉(zhuǎn)移率及轉(zhuǎn)移量的差異進(jìn)行顯著性檢驗(yàn),顯著水平為P= 0.05。
2 結(jié)果與分析
2.1 不同林齡的活枝、葉和凋落枝、葉的養(yǎng)分含量特征
從圖2和圖3看出,同一林齡時(shí),杉木活枝葉中各養(yǎng)分元素濃度大于凋落枝葉中各養(yǎng)分元素濃度。除7,11年生時(shí)活葉與凋落葉中Ca濃度,16,20,25年生時(shí)活葉與凋落葉中Mg濃度差異不顯著外(P>0.05),其余同一林齡時(shí),活葉和凋落葉中同一養(yǎng)分元素濃度之間差異顯著(P<0.05)。同一林齡時(shí)的活枝與凋落枝的Ca濃度之間,Mg濃度之間差異不顯著(P>0.05),活枝與凋落枝的N濃度之間,P濃度之間,K濃度之間差異顯著(P<0.05)。
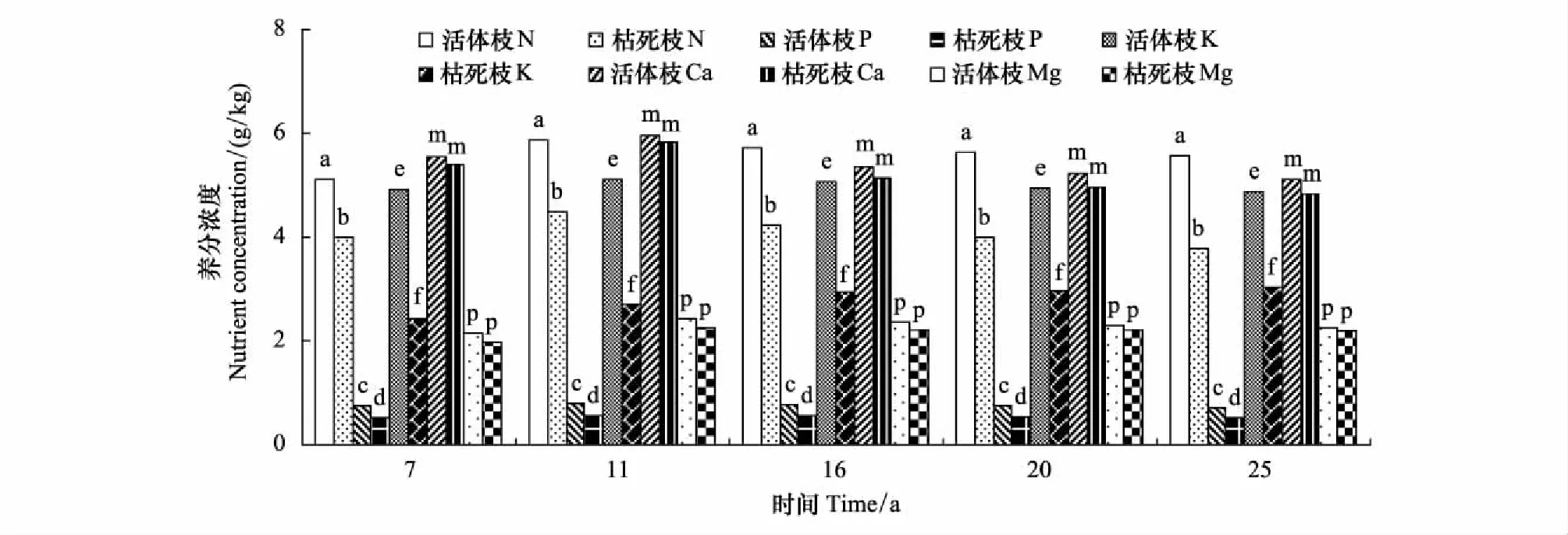
圖2 不同林齡杉木活的和凋落的枝的養(yǎng)分濃度/(g/kg)Fig.2 Nutrient concentrations of live and littering shoots of different ages of Chinese fir同一林齡有相同的小寫字母表示差異不顯著(P>0.05),不相同表示差異顯(P<0.05)
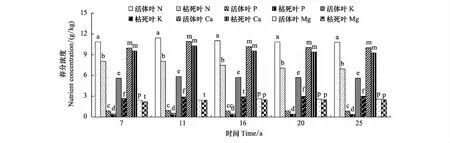
圖3 不同林齡杉木活的和凋落的葉的養(yǎng)分濃度/(g/kg)Fig.3 Nutrient concentrations of live and littering leaves of different ages of Chinese fir同一林齡有相同的小寫字母表示差異不顯著(P>0.05),不相同表示差異顯(P<0.05)
活枝葉與凋落枝葉的各養(yǎng)分元素濃度絕對差值(活枝(葉)與凋落枝(葉)某養(yǎng)分元素的濃度差)大小順序,任一林齡時(shí),葉是N>K>Ca>P>Mg,枝為K>N>Ca>P>Mg;相對差值(活枝(葉)與凋落枝(葉)某養(yǎng)分元素的濃度差與活枝(葉)某養(yǎng)分元素濃度比值)的大小順序,任一林齡時(shí),枝是K>P>N>Mg>Ca;葉則為16年生以前K>P>N>Mg>Ca,16年生后P>K>N>Mg>Ca。
2.2 不同林齡的枝葉凋落量
將每個(gè)林齡段內(nèi)每年測定的凋落枝葉量進(jìn)行年平均統(tǒng)計(jì)得出,會(huì)同杉木林在1—7,8—11,12—16,17—20,21—25年生林齡段,年均凋落的枝葉量依次為0.496,1.730,3.098,4.903和4.694 t hm-2a-1(表2)。不同林齡杉木凋落的枝葉與杉木枝葉的生長周期有關(guān)。杉木是常綠樹種,葉一般要生長5年后才枯死[21];根據(jù)我們在會(huì)同的長期觀測,枝條的生長周期在10年左右。所以杉木林從幼林到10年生左右時(shí)凋落的枝葉量不會(huì)很多;隨著林齡的增加,先前生長的枝、葉逐漸枯死其凋落的量也逐漸增多。杉木枝葉的凋落量也與林分冠層結(jié)構(gòu)特征有關(guān)。一般10年生后的杉木林冠層開始郁閉,為了調(diào)節(jié)冠層內(nèi)的光資源就會(huì)出現(xiàn)自然整枝現(xiàn)象。由于該林分一直未進(jìn)行過間伐,10年生后林冠層枝葉密集,自然整枝過程較強(qiáng)烈,所以12—20年生的林齡段中凋落的枝葉量增加較大,進(jìn)行成熟階段林冠層變得稀疏,自然整枝現(xiàn)象減弱,導(dǎo)致凋落的葉量有所減少。

表2 不同林齡時(shí)杉木林年平均凋落的枝葉量/(t hm-2 a-1)
平均值±標(biāo)準(zhǔn)誤,同一行有相同的小寫字母表示差異不顯著(P>0.05),不相同表示差異顯(P<0.05);某林齡時(shí)同一列中有相同的大寫字母表示差異不顯著(P>0.05),不相同表示差異顯著(P<0.05)
2.3 枝葉凋落前養(yǎng)分轉(zhuǎn)移量特征
2.3.1 枝葉凋落前的養(yǎng)分轉(zhuǎn)移量
表3表明,會(huì)同杉木林1年至25年生內(nèi)不同林齡階段枝葉凋落前的年均養(yǎng)分轉(zhuǎn)移量為3.22—31.89 kg hm-2a-1。其中,枝的養(yǎng)分轉(zhuǎn)移量0.18—8.40 kg hm-2a-1,占總量的2.17%—28.69%,葉的養(yǎng)分轉(zhuǎn)移量3.04—24.52 kg hm-2a-1,占總量71.31%—97.83%。 可見,會(huì)同杉木枝葉凋落前的養(yǎng)分轉(zhuǎn)移量絕大部分由葉貢獻(xiàn)。
從表3還看出,枝的N、P、K、Ca、Mg年均轉(zhuǎn)移量分別是0.05—3.57,0.006—0.41,0.11—3.71,0.006—0.57,0.008—0.18 kg hm-2a-1,分別占枝的轉(zhuǎn)移總量的27.78%—42.50%,3.33%—5.30%,44.17%—61.11%,3.33%—6.79%,1.67%—4.45%。葉的N、P、K、Ca、Mg年均轉(zhuǎn)移量分別是1.25—11.98,0.18—1.52,1.30—8.51,0.19—2.11,0.12—0.43 kg hm-2a-1,分別占葉的轉(zhuǎn)移總量的41.12%—49.32%,5.92%—6.42%,33.24%—42.76%,6.25%—9.68%,1.34%—3.95%。總轉(zhuǎn)移量中的N、P、K、Ca、Mg量依次是1.30—14.85,0.186—1.91,1.41—11.98,0.196—2.59,0.128—0.59 kg hm-2a-1,分別占養(yǎng)分總轉(zhuǎn)移量的40.37%—47.38%,5.29%—5.97%,36.37%—43.79%,6.09%—8.85%,1.43%—3.97%。以上分析看出,同林齡杉木枝葉凋落前的轉(zhuǎn)移養(yǎng)分量中主要是N和K這兩種元素的量,占總量83.75%—84.25%, P、Ca、Mg這3種元素轉(zhuǎn)移量只占總量15.75%—16.25%。
枝葉凋落前的各養(yǎng)分元素轉(zhuǎn)移量與枝葉凋落物量、枝葉凋落前某種元素濃度、以及枝葉凋落前、后的某種元素濃度差值(絕對值)有關(guān)。某林齡段凋落的枝葉量對每一個(gè)元素而言是相同的,因而,決定各養(yǎng)分元素轉(zhuǎn)移量是枝葉凋落前該養(yǎng)分元素濃度,以及枝葉凋落前、后的該元素濃度差值。觀察圖2和圖3,相同林齡枝葉凋落前的N和K濃度高,而且枝葉凋落前、后的N和K的濃度差值都較大,因而其轉(zhuǎn)移量也較多。枝葉凋落前的P的濃度很小,且枝葉凋落前、后的P的濃度差絕對值小,導(dǎo)致其轉(zhuǎn)移量少。雖然枝葉凋落前的Ca、Mg都濃度較大,但枝葉凋落前、后它們的濃度差值很少,所以Ca、Mg的轉(zhuǎn)移量也少。
1—7年生內(nèi)枝凋落前的P與Ca,葉凋落前的P與Ca,K與Mg,8—11年生內(nèi)枝凋落前的Ca與Mg的年均轉(zhuǎn)移量差異不顯著(P>0.05),其余同林齡時(shí)枝葉凋落前的各元素轉(zhuǎn)移量差異顯著(P<0.05)。
表3不同林齡杉木林枝葉凋落前的養(yǎng)分年平均轉(zhuǎn)移量/(kg hm-2a-1)
Table3TheAnnualaverageamountofnutrientstransferredfrombeforelitterfallbranchesandleavesofdifferentage-oldChinesefirplantation
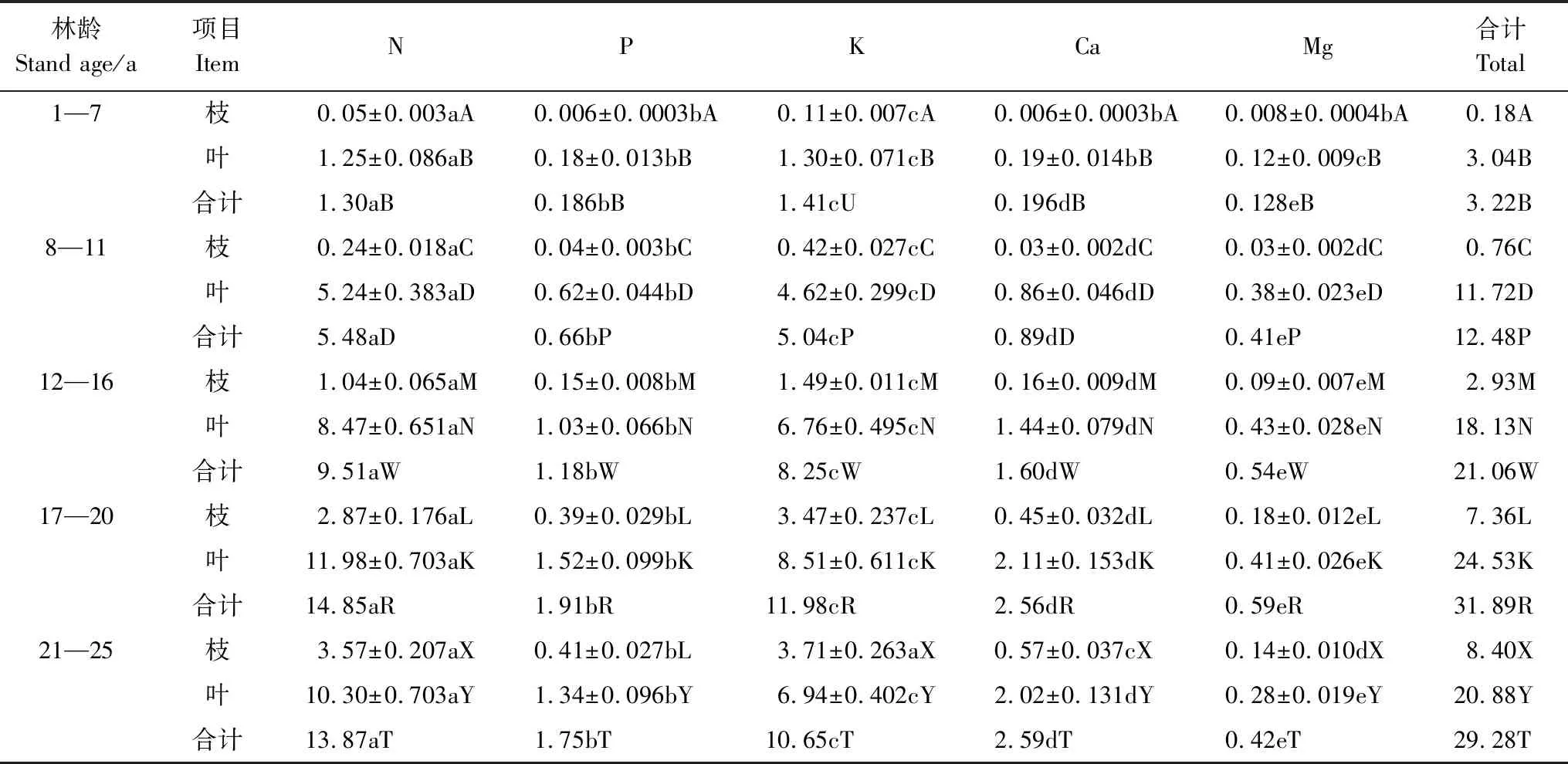
林齡Stand age/a項(xiàng)目ItemNPKCaMg合計(jì)Total1—7枝0.05±0.003aA0.006±0.0003bA0.11±0.007cA0.006±0.0003bA0.008±0.0004bA0.18A葉1.25±0.086aB0.18±0.013bB1.30±0.071cB0.19±0.014bB0.12±0.009cB3.04B合計(jì)1.30aB0.186bB1.41cU0.196dB0.128eB3.22B8—11枝0.24±0.018aC0.04±0.003bC0.42±0.027cC0.03±0.002dC0.03±0.002dC0.76C葉5.24±0.383aD0.62±0.044bD4.62±0.299cD0.86±0.046dD0.38±0.023eD11.72D合計(jì)5.48aD0.66bP5.04cP0.89dD0.41eP12.48P12—16枝1.04±0.065aM0.15±0.008bM1.49±0.011cM0.16±0.009dM0.09±0.007eM2.93M葉8.47±0.651aN1.03±0.066bN6.76±0.495cN1.44±0.079dN0.43±0.028eN18.13N合計(jì)9.51aW1.18bW8.25cW1.60dW0.54eW21.06W17—20枝2.87±0.176aL0.39±0.029bL3.47±0.237cL0.45±0.032dL0.18±0.012eL7.36L葉11.98±0.703aK1.52±0.099bK8.51±0.611cK2.11±0.153dK0.41±0.026eK24.53K合計(jì)14.85aR1.91bR11.98cR2.56dR0.59eR31.89R21—25枝3.57±0.207aX0.41±0.027bL3.71±0.263aX0.57±0.037cX0.14±0.010dX8.40X葉10.30±0.703aY1.34±0.096bY6.94±0.402cY2.02±0.131dY0.28±0.019eY20.88Y合計(jì)13.87aT1.75bT10.65cT2.59dT0.42eT29.28T
平均值±標(biāo)準(zhǔn)誤,同一行有相同的小寫字母表示差異不顯著(P>0.05),不相同表示差異顯(P<0.05);某林齡時(shí)同一列中有相同的大寫字母表示差異不顯著(P>0.05),不相同表示差異顯著(P<0.05)
2.3.2 枝葉凋落前養(yǎng)分轉(zhuǎn)移量的年變化特征
任一林齡段的凋落前的葉中各養(yǎng)分元素轉(zhuǎn)移量大小順序是K>N>Ca>P>Mg。凋落前的枝中轉(zhuǎn)移的各養(yǎng)分元素量大小順序,各林齡段有所不同。1—7年生為K>N>Mg>P = Ca,8—11年生是K>N>P>Ca>Mg,11年生后都為K>N>Ca>P>Mg。從圖2中可知,任一林齡段的活葉中各養(yǎng)分元素濃度大小依次是N>Ca>K>Mg>P。11年生以前活枝中是Ca>N>K>Mg>P,11年生以后是N>Ca>K>Mg>P。將各林齡段中凋落的枝葉中各養(yǎng)分元素轉(zhuǎn)移量大小與各林齡段的活枝葉各養(yǎng)分元素濃度大小相比較,不難看出,凋落的枝葉中某養(yǎng)分元素轉(zhuǎn)移量與活枝葉中這元素濃度相關(guān)性不大。
1—7年生時(shí),枝凋落前的年均養(yǎng)分轉(zhuǎn)移量0.18 kg hm-2a-1,到20—25年生時(shí)上升到8.40 kg hm-2a-1,養(yǎng)分轉(zhuǎn)移量隨林齡增加而增加。在20年生以前,葉凋落前的年均養(yǎng)分轉(zhuǎn)移量不斷上升,20年生以后有所下降,隨林齡的變化呈單峰形曲線。前已分析,枝凋落前的養(yǎng)分轉(zhuǎn)移量占總量的2.17%—28.69%,葉凋落前的轉(zhuǎn)移量占總量71.31%—97.83%。因此,林分枝葉凋落前的養(yǎng)分轉(zhuǎn)移總量隨林齡變化受葉中的養(yǎng)分轉(zhuǎn)移量控制,與葉的年變化規(guī)律相同。細(xì)看表3可知,枝和葉凋落前的N、P、K、Ca轉(zhuǎn)移量的年變化,分別與枝和葉各自的養(yǎng)分轉(zhuǎn)移量的年變化趨勢一樣。Mg有所不同,枝凋落前轉(zhuǎn)移的Mg量,在林分20年生以前呈上升趨勢,20年后呈下降趨勢;葉凋落前轉(zhuǎn)移的Mg量,林分17年生以前不斷增加,17年生后又逐漸下降。
枝葉凋落前的養(yǎng)分轉(zhuǎn)移量隨林齡的變化同樣由不同林齡的凋落枝葉量、枝葉凋落前和后的養(yǎng)分濃度差值決定。將表3和表2比較,不難發(fā)現(xiàn)枝葉凋落前養(yǎng)分轉(zhuǎn)移量的年變化與枝葉凋落量的年變化相吻合。雖然從圖2和圖3可知,枝葉凋落前與后的K與K、Mg與Mg濃度差值隨林齡增加而減少,但K和Mg的轉(zhuǎn)移量仍隨林齡增加而增大。可見,枝葉凋落前養(yǎng)分轉(zhuǎn)移量的年變化主要受不同林齡時(shí)凋落的枝葉量的制約。
17—20年生和21—25年生內(nèi)的枝凋落前的P轉(zhuǎn)移量之間,K轉(zhuǎn)移量之間,以及12—16年生和17—20年生內(nèi)的葉凋落前的Mg轉(zhuǎn)移量之間差異不顯著(P>0.05),其余不同林齡枝或葉凋落前的相同元素轉(zhuǎn)移量差異顯著(P<0.05)(表3)
2.3.3 枝與葉凋落前養(yǎng)分轉(zhuǎn)移量的比較
將枝和葉凋落前的年均養(yǎng)分轉(zhuǎn)移量進(jìn)行比較得出。同一林齡,葉凋落前的N、P、K、Ca、Mg的轉(zhuǎn)移量分別是枝凋落前轉(zhuǎn)移的相應(yīng)元素量的2.9—25,3.2—30,1.9—13,3.5—32,2.0—15倍,且差異顯著(P>0.05)。葉年均養(yǎng)分轉(zhuǎn)移總量是枝的2.5—17倍。這是由于枝凋落前后養(yǎng)分濃度差值大小與葉凋落前后的不同,以及不同林齡時(shí)枝葉凋落量不同造成的。仔細(xì)比較圖2與圖3,同一林齡的葉凋落前后各養(yǎng)分元素濃度差值都大于枝凋落前后的,同林齡時(shí)葉凋落量是枝凋落量的1.3—8.9倍(表2),這就造成葉年均養(yǎng)分轉(zhuǎn)移總量大于枝的,且它們之間差異顯著(P<0.05)。
枝葉凋落前的養(yǎng)分轉(zhuǎn)移量隨林齡的變化也存在差別。枝的養(yǎng)分轉(zhuǎn)移量隨林齡增加而增加,葉隨林齡的變化呈單峰形的曲線,波峰出現(xiàn)在17—20年生內(nèi)。這是因?yàn)橹Φ纳L周期較長,幼林時(shí),枝的凋落量非常少,隨林齡增加,枝的凋落量也逐漸增加。葉的生長周期比樹枝的短,而且前面已分析,由于該林分一直未進(jìn)行過間伐,林冠層枝葉密集,葉生長空間受到限制,所以林分郁閉后到20年生以前凋落的葉量增加較大;進(jìn)行成熟階段林冠層變得稀疏,葉的生長空間得到改善導(dǎo)致凋落的葉量有所減少。正因如此,導(dǎo)致了枝與葉在養(yǎng)分轉(zhuǎn)移量以及隨林齡的變化差別。
2.4 枝、葉凋落前的養(yǎng)分轉(zhuǎn)移率
在1—25年生的任一林齡段,枝凋落前各營養(yǎng)元素轉(zhuǎn)移率的大小順序是K>P>N>Mg>Ca。17—20年生及21—25年生的同一林齡段,枝凋落前的P和N轉(zhuǎn)移率差異不顯著(P>0.05),其余,相同林齡時(shí),各元素轉(zhuǎn)移率之間差異顯著(P<0.05)(圖4)。不同林齡段,葉凋落前各營養(yǎng)元素轉(zhuǎn)移率的大小順序不完全相同,16年生以前葉是K>P>N>Mg>Ca,16年生后為P>K>N>Ca>Mg。12—16年生時(shí),葉凋落前的P和K轉(zhuǎn)移率差異不顯著(P>0.05),其余相同林齡時(shí)葉凋落前的各元素轉(zhuǎn)移率之間差異顯著(P<0.05) (圖5)。
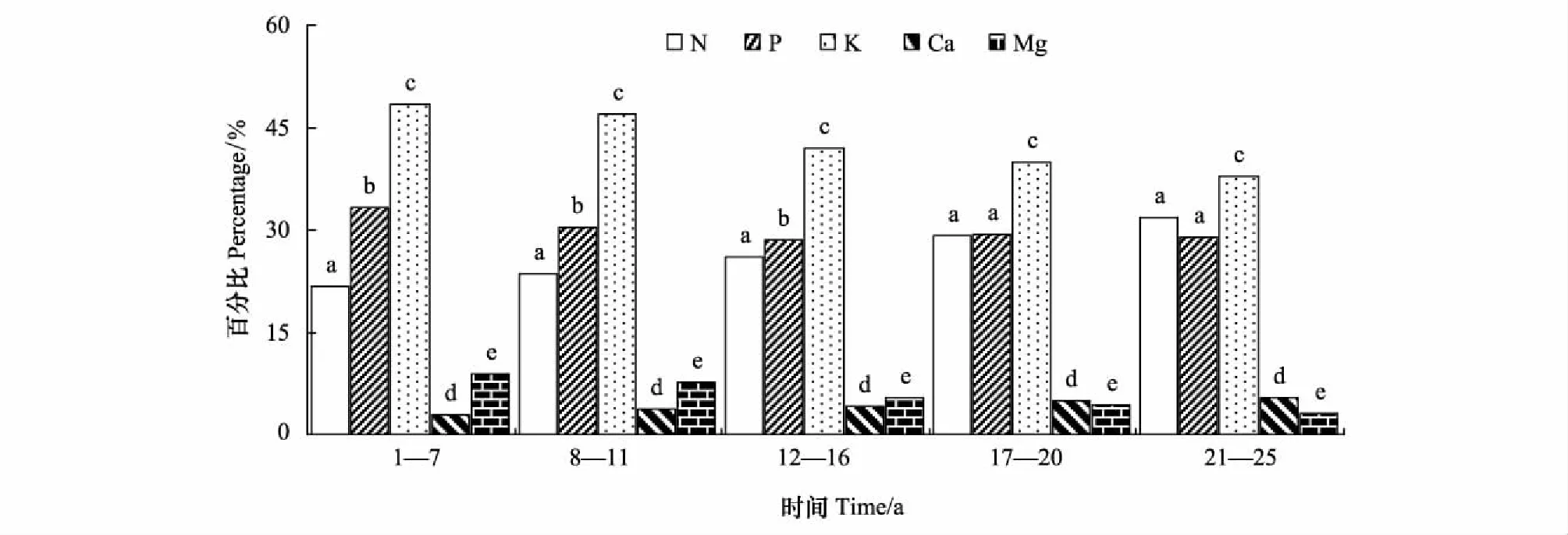
圖4 不同林齡杉木枝凋落前的養(yǎng)分年平均的轉(zhuǎn)移率Fig.4 The average annual reflux rate return of nutrients in branches of Chinese fir plantations in different ages同一林齡有相同的小寫字母表示差異不顯著(P>0.05),不相同表示差異顯(P<0.05)
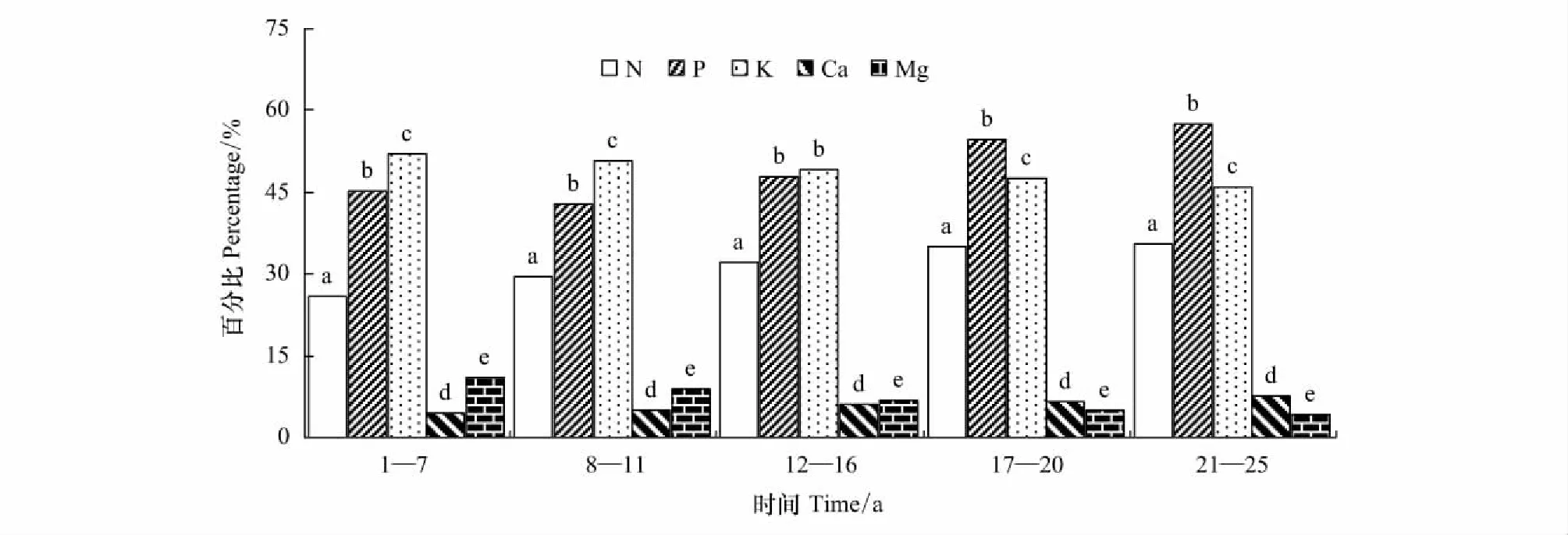
圖5 不同林齡杉木林葉凋落前的養(yǎng)分年平均的轉(zhuǎn)移率Fig.5 The average annual reflux rate return of nutrients in leaves of Chinese fir plantations in different ages同一林齡有相同的小寫字母表示差異不顯著(P>0.05),不相同表示差異顯(P<0.05)
1—7,8—11,12—16,17—20年生和21—25年生林齡段,枝凋落前養(yǎng)分轉(zhuǎn)移率依次為20.97%,21.64%,21.83%,22.27%和22.59%;在上面相應(yīng)林齡段,葉凋落前養(yǎng)分轉(zhuǎn)移率分別是22.98%,24.69%,25.63%,25.87%和26.06%。同一林齡時(shí),葉凋落前的N、P、K、Ca、Mg轉(zhuǎn)移率,分別比樹枝凋落前的相應(yīng)元素轉(zhuǎn)移率大11.20%—25.36%,35.34%—98.27%,7.47%—21.52%,34.00%—52.65%,15.94%—32.59%,且差異顯著(P<0.05)。
枝和葉凋落前養(yǎng)分轉(zhuǎn)移率都隨林齡增加而增大。各林齡段的枝凋落前的養(yǎng)分轉(zhuǎn)移率差異不顯著(P>0.05); 1—7年生內(nèi)葉凋落前的養(yǎng)分轉(zhuǎn)移率與其他林齡段的葉的差異顯著(P<0.05),其余各林齡段之間的葉的養(yǎng)分轉(zhuǎn)移率差異不顯著(P>0.05)。各營養(yǎng)元素轉(zhuǎn)移率隨林齡變化中,枝和葉中稍有差別,枝凋落前的N和Ca轉(zhuǎn)移率隨林齡增加而上升,K、P和Mg隨林齡增加而下降。葉凋落前P轉(zhuǎn)移率年變化與枝相反,但其余元素轉(zhuǎn)移率年變化與枝相同。
植物枝葉凋落前養(yǎng)分向活體組織轉(zhuǎn)移使得養(yǎng)分在植物體內(nèi)被植物再利用,也就是說枝葉具有對養(yǎng)分再吸收利用的能力。養(yǎng)分再吸收利用能力的大小取決于枝葉凋落前的養(yǎng)分轉(zhuǎn)移率,轉(zhuǎn)移率越大其能力越大,反之則然。從會(huì)同杉木枝葉凋落前養(yǎng)分轉(zhuǎn)移率及隨林齡變化趨勢看出,枝葉對養(yǎng)分再吸收利用能力隨林齡增加而增強(qiáng),葉對養(yǎng)分再吸收利用能力大于枝。就具體養(yǎng)分元素而言,枝對K的再吸收利用能力>P>N>Mg>Ca,枝對N和Ca再吸收利用能力隨林齡增加而增加,對K、P和Mg則隨林齡增加而下降。16年生以前,葉對K再吸收利用能力>P>N>Mg>Ca,16年生后則為P>K>N>Mg>Ca,葉對N、Ca 和P再吸收利用能力隨林齡增加而上升,對K和Mg則隨林齡增加而降低。
3 討論
本研究得出,會(huì)同杉木枝、葉凋落前N、P、K向活體的轉(zhuǎn)移量大于Ca、Mg的轉(zhuǎn)移量,這與大部分研究結(jié)果相同[10,22- 25]。會(huì)同杉木葉凋落前各元素的轉(zhuǎn)移率都比刺槐(Robiniapseucdoacacia)[23]、楊樹 (Populus)[24]、樟子松(Pinussylvestrisvar.mongolica)[12]、新銀合歡(Leucaenaleucocephala)[8]、火炬松(PinustaedaL.)[25]低(這些都只是進(jìn)行葉衰老時(shí)養(yǎng)分轉(zhuǎn)移的研究);與福建杉木[13]相比,除葉的Mg相近外,其余枝葉中各元素轉(zhuǎn)移率均低于福建。會(huì)同杉木葉凋落前元素轉(zhuǎn)移率與林寶平等研究的相比[10],P、Ca轉(zhuǎn)移率高于林寶平研究的,N、K、Mg則低于他們。會(huì)同不同林齡時(shí)杉木葉的養(yǎng)分元素轉(zhuǎn)移率,17年生以前K>P>N>Mg>Ca,17年生以后是P>K>N>Mg>Ca,與不同林齡時(shí)樟子松葉(P>K>N>Ca>Mg)[12]、福建杉木葉(P>K>N>Mg>Ca)[13]相比存在差別;會(huì)同杉木枝的養(yǎng)分元素轉(zhuǎn)移率K>P>N>Mg>Ca,也與福建杉木枝[13](P>K>N>Ca>Mg)略有不同。1—25年生會(huì)同杉木葉中N、P、Ca轉(zhuǎn)移率隨林齡增加而上升,K、Mg隨林齡增加而下降,枝中養(yǎng)分元素轉(zhuǎn)移率除P與葉的相反外,其余的與葉的相同,這與10—22年生福建杉木葉和枝的研究結(jié)果一樣[13];與樟子松葉相比,雖然K的年變化一樣,但與其的N和P隨林齡增大而下降[12]的結(jié)論相反。
本研究與同類研究結(jié)果差別是由以下原因產(chǎn)生的。(1)本研究的實(shí)際是枝葉凋落前養(yǎng)分的轉(zhuǎn)移,其他的研究是枝葉枯死前的養(yǎng)分轉(zhuǎn)移。由于枯死枝葉的宿存性,不會(huì)立即從樹上掉落。殘存在樹上枯死枝葉在掉落以前,由于雨水淋溶及風(fēng)化作用仍可能有養(yǎng)分流失到林地,本研究以活體枝葉與凋落后枝葉的養(yǎng)分濃度差估算其養(yǎng)分向活體組級的轉(zhuǎn)移量,有可能高估了其養(yǎng)分轉(zhuǎn)移量。(2)研究的植物種類不同。刺槐、楊樹屬于落葉闊葉樹,新銀合歡是常綠小喬木,黃山松、樟子松、火炬松和杉木則為常綠針葉樹。植物種類不同其生理功能和枝、葉的生長周期不一樣。枝、葉的生長周期不一樣,導(dǎo)致不同林齡段的枝、葉枯死量或凋落量及其養(yǎng)分轉(zhuǎn)移量不一樣。而且,常綠樹種葉壽命較長,能長時(shí)間保持更多的N和P元素,相對而言,落葉樹種葉片中N和P元素濃度較低,因而N和P的重吸收率高于常綠樹種[24- 25]。(3)本研究是在同一林分不同林齡進(jìn)行的,其他同類研究大都用“空間換時(shí)間”法,盡管選擇的是立地條件基本相似,但仍存在某些差別,都有可能導(dǎo)致其研究結(jié)果偏移實(shí)際。(4)本研究把1—25年生林分分成幾個(gè)連續(xù)的林齡段,依據(jù)每林齡段前后測定數(shù)據(jù)分段逐一分析,同類研究則是在林分某一林齡時(shí)測定的數(shù)據(jù),把林分1年生到這個(gè)林齡時(shí)作為一個(gè)時(shí)間序列處理(由于用“空間換時(shí)間”法,沒有該林分以前的數(shù)據(jù)),所取的時(shí)間尺度不一樣,導(dǎo)致研究結(jié)果也不一樣。
從本研究結(jié)果看出,枝葉凋落前的養(yǎng)分轉(zhuǎn)移量除受枝葉凋落前和后的養(yǎng)分濃度差值影響外,更多地受枝葉凋落量的控制。眾所周知,不同的樹種的枝葉生長周期不同,導(dǎo)致不同林齡時(shí)凋落的枝葉量不一樣;同一樹種不同生育階段生長的枝葉量及林冠層枝葉密集度不同,因此不同生育階段的枝葉凋落量也可能不相同。這些都能影響?zhàn)B分轉(zhuǎn)移量的大小和年變化特征。從這點(diǎn)上分析,樹木枝、葉凋落前養(yǎng)分轉(zhuǎn)移量及其年變化與樹木的生理特性、生長發(fā)育特征有很大的關(guān)聯(lián)。
本研究7年生后各林齡段的葉凋落前養(yǎng)分轉(zhuǎn)移率差異不顯著(P>0.05)。這與林寶平等[10]研究的相同(研究的是8、14、21年生2代杉木林和46年生1代杉木林)。表明對杉木而言,葉枯死養(yǎng)分轉(zhuǎn)移率的大小與杉木生長年齡關(guān)系不大。已有研究表明,常綠闊葉林葉的N、P的轉(zhuǎn)移率比落葉闊葉林的都低[24- 25]。這說明植物枝葉凋落前養(yǎng)分轉(zhuǎn)移率與植物的生活型有關(guān)。會(huì)同不同林齡段杉木林枝葉凋落前的養(yǎng)分轉(zhuǎn)移量中,枝的占總量的5.59%—28.69%,葉占71.31%—94.41%,可見,葉在養(yǎng)分內(nèi)部轉(zhuǎn)移中起主要作用。這與費(fèi)世民等[25]研究的結(jié)果相同。而且也證實(shí)了Miller等[28]認(rèn)為葉是林木體內(nèi)養(yǎng)分轉(zhuǎn)移的最重要的源的結(jié)論。
從本研究的結(jié)果看出,不僅葉枯死前可以發(fā)生養(yǎng)分轉(zhuǎn)移,枝枯死前也可以發(fā)生養(yǎng)分轉(zhuǎn)移,而且不同粗細(xì)、不同林齡的枝條在枯死前都存在養(yǎng)分的遷移。這點(diǎn)在福建三明市的杉木的同類研究中[13]得到證實(shí)。用“時(shí)空轉(zhuǎn)換法”研究的福建三明市幼齡林(10年生)、成齡林(22年生)、過熟林(34年生)杉木養(yǎng)分內(nèi)循環(huán)發(fā)現(xiàn),不同林齡時(shí)的杉木活枝的養(yǎng)分濃度均極顯著高于枯枝的養(yǎng)分濃度[13]。這實(shí)際意味著所有枝條枯死前已發(fā)生養(yǎng)分轉(zhuǎn)換。
本研究估算不同林齡段的杉木枝葉的養(yǎng)分轉(zhuǎn)移量,并不是以某林齡段內(nèi)枯死的枝葉量估算的,而是以某林齡段內(nèi)脫離樹體(凋落)的枝葉量估算的。由于枯死枝葉的宿存性,枯死枝葉不會(huì)立即從樹上掉落,有些還殘存在樹上。本文并沒有估算這些存留在樹上的枯死枝葉的養(yǎng)分轉(zhuǎn)移量,因此,本文研究枝葉的養(yǎng)分轉(zhuǎn)移量比實(shí)際的養(yǎng)分轉(zhuǎn)移量偏低。如何估算某林齡宿存的枯死枝葉量,目前還沒有較可行的方法。如果,得知不同林齡時(shí)的枝葉生物量和枝葉的生長周期,用枝葉生長周期估算不同林齡時(shí)的枝葉枯死量,就能較好地解決這問題。例如,若杉木5年生時(shí)生產(chǎn)的枝葉量分別為W1和W2,枝葉生長周期分別5年和10年。那么林分10年生時(shí),葉的枯死量為W1,林分15年生時(shí),枝的枯死量為W2,而不必管枯死的枝葉是殘存在樹上還是掉落了。這樣推算出的不同林齡時(shí)的枝葉枯死量,用來估算不同林齡時(shí)的枯死枝葉的養(yǎng)分轉(zhuǎn)移量,其結(jié)果更逼近實(shí)際。
目前對植物枝、葉衰老時(shí)養(yǎng)分向活體轉(zhuǎn)移的機(jī)制還不完全了解。雖然不少研究表明,從土壤貧瘠生境中植物養(yǎng)分再吸收效率高于土壤肥沃生境中植物[27-29],也有研究指出植物養(yǎng)分轉(zhuǎn)移與土壤養(yǎng)分供給性大小相關(guān)性不大[30],貧瘠土壤并不一定是植物體內(nèi)養(yǎng)分轉(zhuǎn)移的驅(qū)動(dòng)力。由此看來,植物體內(nèi)養(yǎng)分轉(zhuǎn)移再利用只是植物對養(yǎng)分貧瘠環(huán)境的一種適應(yīng)機(jī)制,也是植物保存養(yǎng)分并維持體內(nèi)養(yǎng)分平衡的一種重要養(yǎng)分利用策略[31]。枝、葉衰老時(shí)養(yǎng)分向活體養(yǎng)分轉(zhuǎn)移的機(jī)理有可能是植物本身具有這種自我調(diào)控的反饋機(jī)制,通過這種調(diào)控機(jī)制使養(yǎng)分再吸收和再利用,這種機(jī)制可能是喬木在自然界長期的生存競爭中形成的[17- 18]。一些研究者用植物生理中“源-匯”的理論分析樹木調(diào)節(jié)體內(nèi)的養(yǎng)分轉(zhuǎn)移的機(jī)制。認(rèn)為樹木器官在某些生理?xiàng)l件下是養(yǎng)分的源,在另一些生理?xiàng)l件下可能是匯,正由于這種匯源的作用,調(diào)控植物枝葉衰老時(shí)養(yǎng)分向活體流動(dòng)[28]。植物枝、葉衰老時(shí)養(yǎng)分向活體的轉(zhuǎn)移是一個(gè)非常復(fù)雜的生理生化過程,對它們的形成機(jī)制的研究仍是一個(gè)非常薄弱的環(huán)節(jié),揭示體內(nèi)的養(yǎng)分轉(zhuǎn)移內(nèi)在調(diào)控機(jī)制仍然是生理生態(tài)學(xué)亟待解決的重要問題。
4 結(jié)論
會(huì)同杉木林枝葉凋落前年均養(yǎng)分轉(zhuǎn)移量為3.22—31.89 kg hm-2a-1。主要是葉的轉(zhuǎn)移量,占71.31%—94.41%,枝只占5.59%—28.69%。就各養(yǎng)分元素而言,N和K占總量83.75%—84.25%,P、Ca、Mg只占總量15.75%—16.25%。
枝的養(yǎng)分轉(zhuǎn)移量隨林齡增加而增加,葉的養(yǎng)分轉(zhuǎn)移量從幼林到近熟林階段逐漸上升,成熟林階段開始下降。養(yǎng)分轉(zhuǎn)移總量隨林齡變化除受不同林齡段枝葉凋落前和后的養(yǎng)分元素濃度差值影響外,更多地受不同林齡段的枝葉凋落量控制。
任一個(gè)林齡階段,枝凋落前各養(yǎng)分元素轉(zhuǎn)移率的大小依次都是K>P>N>Mg>Ca;葉凋落前各元素轉(zhuǎn)移率的大小順序是,林分16年生以前,K>P>N>Mg>Ca,16年生后,P>K>N>Mg>Ca。枝和葉的養(yǎng)分轉(zhuǎn)移率都隨林齡增加而增大,各林齡段的枝凋落前的養(yǎng)分轉(zhuǎn)移率差異不顯著(P>0.05), 1—7年生內(nèi)葉凋落前的養(yǎng)分轉(zhuǎn)移率與其他林齡段的葉的差異顯著(P<0.05),其余各林齡段之間的養(yǎng)分轉(zhuǎn)移率差異不顯著(P>0.05)。
會(huì)同杉木枝葉對養(yǎng)分再吸收利用能力隨林齡增加而增強(qiáng),葉的養(yǎng)分再吸收利用能力大于枝。就具體養(yǎng)分元素而言,枝對K的再吸收利用能力>P>N>Mg>Ca;對N和Ca再吸收利用能力隨林齡增加而增加,對K、P和Mg則隨林齡增加而下降。16年生以前葉對K再吸收利用能力>P>N>Mg>Ca,16年生后則為P>K>N>Mg>Ca;對N、Ca 和P的再吸收利用能力隨林齡增加而上升,K和Mg則隨林齡增加而降低。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻(xiàn)通報(bào)(2022年3期)2022-05-23 13:46:54
天津外國語大學(xué)學(xué)報(bào)(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
民用飛機(jī)設(shè)計(jì)與研究(2019年4期)2019-05-21 07:21:24
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
汽車工程學(xué)報(bào)(2017年2期)2017-07-05 08:13:02

- 生態(tài)學(xué)報(bào)的其它文章
- 棉稈炭特性及其對灰漠土土壤有機(jī)碳礦化的效應(yīng)
- 央地兩級政府生態(tài)治理行動(dòng)的演化博弈分析——基于財(cái)政分權(quán)視角
- 基于擴(kuò)展耗模型的可持續(xù)發(fā)展水平區(qū)域空間差異——以中國31個(gè)省市為數(shù)據(jù)源
- 新薛河溪流生境底棲動(dòng)物群落結(jié)構(gòu)季節(jié)動(dòng)態(tài)
- 北方農(nóng)牧交錯(cuò)帶退耕還林與還草對地面節(jié)肢動(dòng)物群落結(jié)構(gòu)的影響
- 新疆天山森林公園土壤螨類群落多樣性與環(huán)境因子的相關(guān)性