豬MII期卵母細胞冷凍前后全基因組表達譜分析
2019-05-05 01:18:00戴建軍吳彩鳳張樹山孫玲偉陳亞寧張德福
上海農業學報 2019年2期
戴建軍,吳彩鳳,張樹山,孫玲偉,陳亞寧,張德福*
(1上海市農業科學院畜牧獸醫研究所,上海 201106;2上海市農業遺傳育種重點實驗室動物遺傳工程研究室,上海 201106;3上海種豬工程技術研究中心,上海 201302)
繼1972年Whittingham等[1]首次報道冷凍小鼠胚胎并獲得存活后代以來,冷凍保存技術已經在超過20個物種中得到了廣泛的應用。然而不同的物種之間,其胚胎或卵母細胞的抗凍能力不一致。在牛、羊上,其玻璃化冷凍保存已經獲得了較大范圍的推廣應用,然而豬卵母細胞和胚胎的冷凍效率至今仍較低[2],冷凍損傷機理研究十分迫切。
玻璃化冷凍可對卵母細胞或胚胎造成超微結構[3]、細胞器功能[4]和相關功能基因表達[5-6]的影響,目前常用電子顯微鏡、共聚焦顯微鏡和RT-PCR等方法進行檢測。然而細胞的生命活動是一個非常復雜的生理生化過程,很難通過這類常規檢測對整個生理生化過程進行系統評估。基因組表達譜芯片具有靈敏、高效、平行化、高通量等優點,通過篩選特異性表達基因,可在基因水平上揭示胚胎發育過程的本質,并在胚胎工程研究中被廣泛應用。目前將芯片技術應用在胚胎方面的研究物種包括人、鼠、牛、豬和山羊等[7]。
目前,已見小鼠[8]、兔[9]和牛[10]的冷凍胚胎的全基因組表達譜分析的相關報道,但未見豬卵母細胞冷凍前后全基因組表達譜分析的相關研究。本研究擬利用Affymetrix豬的全基因組表達譜芯片,對玻璃化冷凍前后的豬MII期卵母細胞進行全基因組表達譜分析,為深入探究其冷凍損傷機理奠定基礎,豐富其低溫生物學的研究內容。
1 材料與方法
除特別說明外,本研究所用試劑均購自美國Sigma公司。
1.1 卵母細胞的采集和體外成熟
于上海五豐公司屠宰場采集初情期前的豬卵巢,置于含500 IUmL雙抗的37 ℃生理鹽水中,2 h內運回實驗室。用18號針頭注射器收集卵丘卵母細胞復合體(COCs),沉淀30 min后取沉淀物,在顯微鏡下挑選有3層以上顆粒細胞且胞質均勻的COCs,經臺氏液和TCM-199(Gibco,美國)洗滌后置于預溫平衡的體外成熟培養液(IVM)中培養。體外成熟培養液配方為:TCM-199中分別添加10%(vv)胎牛血清(Gibco,美國)、10%(vv)豬卵泡液(實驗室自制)、10 IUmL孕馬血清促性腺激素(寧波市第三激素制品有限公司,中國)、10 IUmL人絨毛膜促性腺激素(寧波市第三激素制品有限公司,中國)、100 IUmL雙抗、0.1 mgmL的L-半胱氨酸和10 ngmL表皮生長因子。培養條件為:38.5 ℃,5% CO2濃度,飽和濕度。培養44 h后的COCs用0.1%的透明質酸酶反復吹打和消化,在顯微鏡下挑出排出第一極體的卵母細胞用于后續試驗。同一批次卵母細胞均隨機分為新鮮組和OPS玻璃化冷凍組進行試驗。
1.2 OPS玻璃化冷凍與解凍
在39℃的恒溫操作臺上,MII期卵母細胞在冷凍保護劑Ⅰ(TCM199中含7.5%的乙二醇,7.5%二甲基亞砜和10%胎牛血清)中處理5 min,然后轉入冷凍保護劑Ⅱ(TCM199中含17%乙二醇,17%二甲基亞砜,10%胎牛血清和0.4 molL蔗糖)中處理15 s,裝入OPS管,30 s內投入液氮保存。解凍時,分別在0.5 mmolL和0.25 mmolL的蔗糖中平衡5 min,然后移入TCM199中洗2遍,備用。冷凍卵母細胞在含10% FBS的TCM199中孵育2 h后進行收集,每150枚卵母細胞為一個重復樣本,單獨保存于-80℃冰箱中用于RNA提取。新鮮組則挑取同一批次150枚新鮮成熟卵母細胞,于-80℃保存后用于RNA提取。
1.3 凍后卵母細胞存活和孤雌激活發育能力檢測
1.4 卵母細胞總RNA的提取
-80℃保存的新鮮和冷凍卵母細胞用于總RNA提取,采用 RNApreo Pure微量樣品總RNA提取試劑盒(TIANGEN)進行RNA提取,用微孔板分光光度計進行RNA濃度檢測,選取A260A280 在1.9—2.1,質量濃度大于5 ngμL的RNA溶液用于總RNA的整體擴增。整體擴增使用TargetAmpTM2-Round aRNA Amplification Kit 2.0試劑盒進行。
1.5 芯片雜交、掃描和數據分析
新鮮組和OPS玻璃化冷凍組分別取3個重復,每個樣本分別取100 ng的上述總RNA,利用RNeasy MinElute Cleanup Kit(Qiagen)純化得到mRNA。目的RNA的準備和芯片雜交、掃描和初步分析處理參照GeneChip?3’ IVT Expres Kits的標準流程進行。GeneChip?Porcine Genome Array來源于Affymetrix公司。
采用Affymetrix 公司提供的Expression Console軟件進行Robust Multichip Analysis(RMA)分析,根據樣品分組,進行組間統計學ANOVA比對分析,得到組間大于2倍差異表達的基因,篩選出新鮮組和OPS玻璃化冷凍組差異表達基因。
對差異表達基因進行顯著性富集分析(Gene ontology,GO)和KEGG Pathway分析。
1.6 部分差異表達基因的qRT-PCR驗證
冷凍和新鮮卵母細胞經RNA提取和反轉錄,應用實時熒光定量PCR技術(qRT-PCR)在代表性基因中隨機選擇3個上調和3個下調表達基因(激酶和轉移酶活性相關基因MAP3K7,細胞骨架相關基因ARPC2和MYL6,脂多糖結合蛋白相關基因LBP和線粒體功能相關基因COX6C、POLG)進行表達驗證。引物信息如表1所示。反應體系:cDNA 模板2.0 μL、SYBR Premix Ex Taq II 10 μL、Forward Primer 0.8 μL、Reverse Primer 0.8 μL、ROX II 0.4 μL、加RNase-Free ddH2O補至20 μL。反應條件:預變性95℃ 30 s,變性95℃ 5 s,退火34 s(Tm均為55℃),72℃延伸30 s,循環35次,退火處收集熒光。在ABI 7500實時熒光定量PCR 儀上運行。冷凍和新鮮卵母細胞各選擇3個樣本,每個樣品重復3次。

表1 RT-PCR 引物信息
1.7 生物統計
qRT-PCR以GAPDH為內參基因,新鮮組和冷凍組各基因的相對表達量采用2-ΔΔCt法計算比較。卵母細胞發育能力采用SPSS 16.0統計軟件進行統計分析,P<0.05表示差異顯著。
2 結果與分析
2.1 冷凍對豬MII期卵母細胞存活和體外發育能力的影響
玻璃化冷凍對豬MII期卵母細胞存活率和孤雌發育能力的影響見表2。OPS玻璃化冷凍后其FDA染色存活率(72.50%)顯著低于新鮮組(98.00%)(P<0.05)。OPS冷凍卵母細胞解凍后經孤雌激活,取得了14.56%的卵裂率和4.85%的囊胚率,顯著低于新鮮組的80.91%和35.45%(P<0.05)。

表2 冷凍對豬卵母細胞存活和孤雌激活發育的影響
同列不同字母表示差異顯著(P<0.05)
2.2 冷凍卵母細胞全基因組表達譜芯片研究
將OPS玻璃化冷凍組數據與新鮮卵母細胞組數據進行比較,篩選出171個2倍以上差異表達基因,其中上調表達基因103個,下調表達基因68個。部分代表性差異表達基因見表3。
2.3 部分差異基因的qRT-PCR表達驗證
如圖1所示,與新鮮卵母細胞相比,MAP3K7 、ARPC2和LBP基因的表達水平在冷凍卵母細胞中分別上調2.98倍、1.92倍和2.65倍,COX6C、POLG和MYL6基因的表達水平分別下調5.64倍、3.26倍和4.85倍,所得結果與基因芯片基本一致。
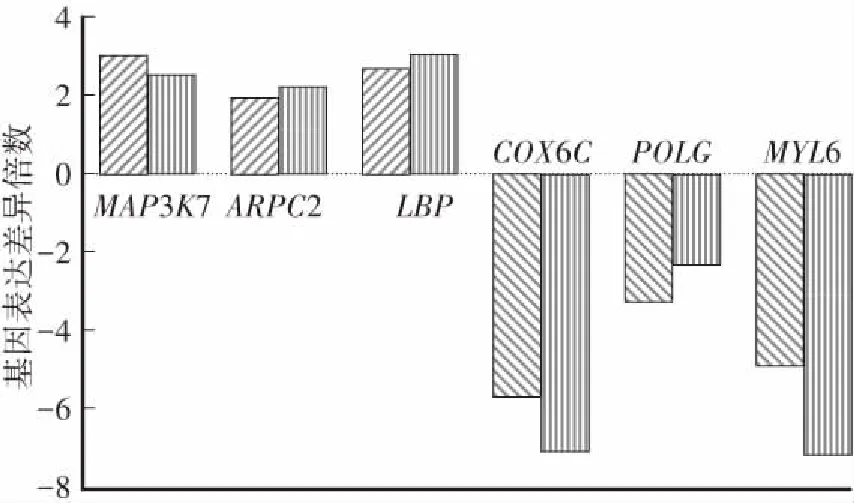
圖1 差異表達基因的qRT-PCR驗證Fig.1 qRT-PCR validation of differentially expressed genes
2.4 芯片結果的系統聚類分析和主成分分析
如圖2所示,3個新鮮組樣品和3個玻璃化冷凍組樣品組內相似性較好,而組間的基因表達具有一定的差異性。
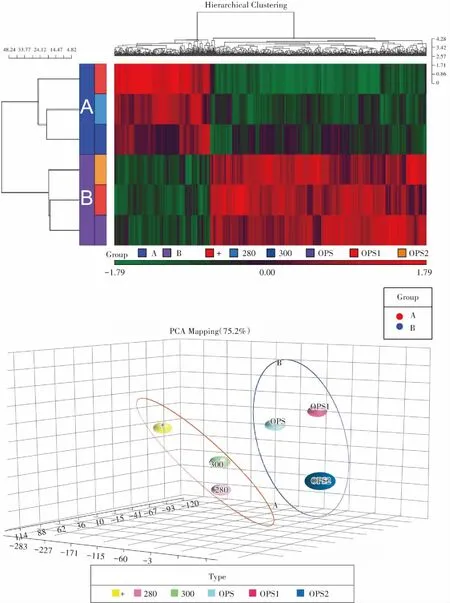
圖2 系統聚類分析和主成分分析Fig.2 Hierarchical clustering analysis and principle component analysis
2.5 冷凍后上調差異表達基因的GO分析
卵母細胞冷凍后上調差異表達基因的生物過程(Biological process,BP)、細胞組成(Cellular component,CC)和分子功能(Molecular function,MF)的GO分析結果見圖3。結果顯示,上調表達基因涉及損傷應答、有機物反應、蛋白激酶調節、細胞增殖、細胞凋亡和細胞壞死調節等生物過程。同時還涉及胞外區、基底質膜、細胞外間隙、質膜、肌動蛋白細胞骨架等細胞組成調控和類固醇結合、酶抑制劑的活性、蛋白質復合物結合、激素結合和轉錄因子活性等分子功能調控。

圖3 上調差異表達基因的GO注釋Fig.3 GO annotation of up-regulated differentially expressed genes
2.6 冷凍后下調差異表達基因的GO分析
卵母細胞冷凍后下調差異表達基因的生物過程、細胞組成和分子功能的GO分析結果見圖4。結果顯示,下調表達基因涉及RNA拼接,核mRNA拼接,mRNA 代謝,轉運正向調節和細胞增殖等生物過程。同時還涉及線粒體脊、線粒體、細胞器膜、線粒體包膜和剪接體等細胞組成調控和核苷酸結合、FAD結合、輔酶結合、固醇轉運活性、核苷酸轉移酶活性和ATP酶活性等分子功能調控。

圖4 下調差異表達基因的GO注釋Fig.4 GO annotation of down-regulated differentially expressed genes
2.7 冷凍后差異表達基因的KEGG Pathway分析
豬MII期卵母細胞玻璃化冷凍后有顯著性差異的KEGG Pathway分析結果見圖5。差異表達基因涉及Toll樣受體信號通路、NOD樣受體信號通路、黏著連接和MAPK信號通路等。

圖5 差異表達基因KEGG Pathway—(-LgP)柱狀圖Fig.5 Histogram of KEGG Pathway—(-LgP)for differentially expressed genes
3 討論
卵母細胞和胚胎的發育過程是由基因控制的復雜過程,然冷凍可造成其基因表達水平的異常變化[5,8]。本研究利用Affymetrix豬商業化表達譜芯片對冷凍前后的豬卵母細胞的全基因組表達譜進行了研究,獲得了已獲注解的2倍以上差異表達基因171個,這為今后篩選得到更多的冷凍相關基因提供了豐富的基礎數據。
在這些差異表達基因中,凍后下調倍數最高的是SLC25A3基因,達38.583倍,該基因是編碼Pic基因的重要組成部分,而Pic則是線粒體內膜上的一個重要功能蛋白,在線粒體膜通道孔打開中發揮重要作用[11]。在文中所列的其他差異表達基因中,ATP2A2、VDAC2、COX6C、POLG、PPARGC-1等均與線粒體功能密切相關。研究表明,冷凍可造成豬MII期卵母細胞線粒體功能的系統性損傷,包括超微結構異常、膜電位下降、ATP水平降低和抗氧化能力減弱等[4]與本研究結果一致。本研究中上調倍數最高的基因為NPPC,上調倍數為3.64317。該基因在抑制卵母細胞的減數分裂中發揮重要作用[12],驗證了本研究中凍后發育能力顯著下降這一結果。
在上調差異表達基因的生物學過程GO分析中,排名第一的為損傷應答(Responese to wounding)。冷凍作為一種強刺激源,使細胞受到嚴重的破壞和損傷,從而使細胞對損傷做出迅速的反應,這為日后冷凍損傷的機理研究提供了新的方向。本研究中凋亡、細胞程序性死亡和細胞壞死調節也是參與基因上調表達的重要生物學過程。研究表明,在卵母細胞和胚胎冷凍中,細胞凋亡被認為是發育能力下降的主要因素之一[13-14],且死亡受體介導的外源性凋亡途徑和線粒體介導的內源性凋亡途徑共同導致了凋亡的發生[15]。此外,有機物反應和蛋白激酶調節等生物學過程也參與了凍后卵母細胞的基因調控,相關研究值得后續進一步深入。
在上調基因的細胞組分分析中,胞外區、基底質膜、細胞外間隙、質膜、肌動蛋白細胞骨架等參與了其冷凍后上調基因表達調控。冷凍作為一種外源性刺激因子,其首先對細胞外區域產生重大影響,因此胞外區分離和細胞外間隙的細胞組分影響排在細胞組分分析的前列。冷凍能對細胞質膜產生嚴重破壞,這一點已經在眾多的研究中得到了證實[16]。冷凍還對卵母細胞的細胞骨架產生影響[17],本試驗中上調基因的細胞組分分析中,有6個上調表達基因參與了肌動蛋白細胞骨架的調控。
卵母細胞冷凍后下調差異表達基因的生物過程分析中,RNA拼接、核mRNA拼接和mRNA 代謝等參與了冷凍后下調基因表達的生物過程調控。MII期卵母細只能通過其自身儲備的RNA和mRNA來合成相關蛋白,其本身并不能轉錄RNA,故其RNA豐度對隨后的胚胎發育有重要作用[18]。本研究芯片結果表明,卵母細胞玻璃化冷凍一方面通過影響卵母細胞內RNA和mRNA的剪接而影響其生物學過程,另一方面通過干擾其mRNA代謝,降低其自身RNA儲備影響基因表達。在本研究的下調基因細胞組分分析中,與線粒體功能密切的組分占了最主要部分,包括線粒體脊、線粒體、細胞器膜和線粒體包膜等,說明冷凍對線粒體功能產生了巨大影響。已有研究表明,冷凍會對卵母細胞的線粒體功能產生破壞[19],玻璃化冷凍會對豬卵母細胞線粒體超微結構、分布和膜電位產生負面影響[4],這些結果與本研究中下調功能基因的細胞組分分析結果相一致。
在凍后差異表達基因的KEGG Pathway分析中,Toll樣受體信號通路、NOD樣受體信號通路、黏著連接和MAPK信號通路參與了差異表達基因的調控通路。Toll樣受體信號通路和NOD樣受體信號通路在機體免疫系統[20]和抗感染[21]中發揮重要作用,冷凍對細胞的損傷和刺激可能誘導了該信號通路的發生,相關研究需進一步深入。黏著連接是細胞中的一個重要連接方式,是一種有細胞骨架參與的緊密連接方式,與細胞信號轉導密切相關[22]。冷凍中黏著連接上調信號通路的參與可能與細胞骨架相關基因上調表達相關。MAPK(絲裂原活化蛋白激酶)是細胞內的一類絲氨酸/蘇氨酸蛋白激酶。研究證實,MAPK信號轉導通路存在于大多數細胞內,在將細胞外刺激信號轉導至細胞及其核內并引起細胞生物學反應(如細胞增殖、分化、轉化及凋亡等)的過程中具有至關重要的作用[23]。冷凍后,MAPK信號通路調控參與其中,表明冷凍對卵母細胞隨后的存活率和發育能力將產生嚴重的影響。
4 結論
本研究利用豬全基因組表達譜芯片分析了豬MII期卵母細胞冷凍前后的基因表達差異,篩選出了171個差異表達基因。聚類分析和KEGG Pathway分析發現細胞損傷應答、凋亡、細胞壞死、細胞質膜、細胞骨架、mRNA拼接和線粒體功能等多種生物學過程參與了凍后卵母細胞的發育異常。本研究為進一步探究豬卵母細胞冷凍損傷機理提供了基礎數據。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
小天使·一年級語數英綜合(2019年8期)2019-08-27 02:23:00
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
中國特種設備安全(2018年11期)2019-01-08 02:08:32
小學科學(學生版)(2018年7期)2018-08-13 09:33:04
汽車工程學報(2017年2期)2017-07-05 08:13:02
