核桃CBF基因在低溫脅迫中表達
2019-05-05 05:36:10宋靜武
湖北林業科技 2019年2期
宋靜武 彭 磊
(1.秭歸縣林業局 林業科學技術推廣中心 宜昌 443600; 2.云南農業大學 園林園藝學院 昆明 650201)
1 實驗目的
核桃(JuglansregiaL.)又被稱為胡桃、羌桃,是一種胡桃科植物。核桃極具營養價值,核桃中脂肪和蛋白質含量較高,又含有人體所需很多種礦物質和微量元素,又富含多種維生素。核桃是一種極為重要的經濟樹種,能為我國核桃產區帶來極大地經濟效益。核桃樹抗寒能力較弱,在受到低溫凍害、寒害以及晚霜等非生物脅迫時會造成花芽枯死和枝條抽干,這會影響樹體的正常生長結實,會造成嚴重的經濟損失,并限制了良種核桃的地理分布。為了提高核桃的經濟和產業價值,研究人員利用現代基因工程技術和分子生物學方法,希望能夠改良核桃品質、探索核桃抗寒機制的分子機理、運用轉基因技術培育高抗性核桃新品種等工作進行研究。
CBF/DREB1(C-repeat binding factors /dehydration response element binding factors1)蛋白是植物體內低溫信號傳導途徑中研究最為廣泛的轉錄因子基因[1]。這個家族在抗寒,抗堿,抗鹽方面有很大作用[2]。擬南芥作為一種常用模式植物,Yamaguchii[3]在極端的干旱和低溫脅迫的條件下對擬南芥進行研究時,發現擬南芥的RD29A基因的啟動子區域中分離出了了DRE順式作用元件(1994),這些元件含有相同的CCGAC序列。隨著研究發現CRT/DRE和其核心序列,常出現在冷誘導以及干旱誘導基因的啟動子中,這是冷誘導過程中必要的一類基因[4]。2002 年,Haake等又分離出了這個家族的第 4 個成員CBF4,CBF4的轉基因植物能觀測與表達和CBF1一樣具有抗寒性顯著提高結果[5]。氨基酸序列分析表明,CBF中含有AP2結構區,AP2結構域第一個DNA結合區,它的作用是結合CRT/DRE順式作用元件,誘導相關基因表達,通過合成膜穩定蛋白、脯氨酸、糖類等物質,改變細胞生理環境進而提高植物的抗寒性[6]。CBF基因的冷誘導不會受自身調節影響,所以這個轉錄因子應該是其他蛋白,他們假設這種轉錄因子為ICE(inducer of CBF expression)。正常條件下ICE不處于活動狀態,植物體可能通過某種條件將ICE隔離在體內,當植物受到冷刺激時ICE會被釋放出來,從而激活一條信號途徑,使CBF基因能夠進行表達[7]。Chinnusamy等在2003年首次分理出ICE1基因[8],經過數據查詢發現ICE1的C端含有一個近似MYCbHLH(MYC-likebasic helix-loop-helix)結構域,這個結構與bHLH蛋白相似,bHLH蛋白能夠識別DNA中的某些元件,而CBF3中含有多個這樣的元件[9],這也說明了ICE能特異的與CBF3中的特定識別序列結合。LOS4-1是CBF基因的一個正向調控因子,是由Guo研究擬南芥突變體時發現的(2002),這種基因突變的擬南芥對低溫的抵抗能力比野生型在24H內降低了40%[10]。LOS基因編碼一種能夠抑制STZ/ZAT10的活性的生物活性酶(STZ/ZAT10能夠抑制冷誘導基因RD29的表達)[11],但是LOS的一個突變LOS4-2卻會增強CBF及其靶基因的表達[12]。Lee[13]等發現了一個與滲透壓相關的CBF負調節因子稱為HOS1(high expression of osmoticstress regulated gene expression 1),HOS編碼環脂蛋白,當受到低溫刺激時會在細胞核內大量積聚,抑制CBF基因表達。Liu等的研究表明,正常狀況下PtrHOS1基因在枳(Poncirus trifoliata (L.) Raf.)莖葉、根部都有很強的表達,經過低溫處理后出現了短暫的下調,說明HOST1基因表達受到低溫影響[14]。SFR6蛋白(sensitive to freezing 6)是一種對低溫敏感的蛋白也是一個特例,這種蛋白是CBF下游正調節因子,能夠下調那些啟動子結構不含有CRT/DRE元件的基因[15~17]。在植物受到低溫刺激時,CBF基因轉錄水平會迅速提高,隨后含有CRT/DRE調節元件的COR基因啟動子會與CBF基因結合進一步促進COR表達。COR基因在低溫條件系會對基因表達進行調節,會降低細胞內的一部分多余物質的含量,從而提高植物抗寒性[18~19]。COR是一種冷應答蛋白,當植物受到干旱鹽堿寒冷的脅迫時會被誘導產生[20]。目前對于COR類基因的研究較多,如擬南芥的COR78基因啟動子常用于非生物脅迫誘導啟動子來增強植物抗逆性,這種啟動子比組成型啟動子更能減小植物生長中的一些負面作用[21]。COR15a基因表達產生的物質會與葉綠體結合,形成一種能夠保護原生質體和葉綠體的物質,進而提高植物細胞的抗寒性[22]。 有關研究表明,CBF基因為AP2/EREBP 超家族的DREB家族成員,是從擬南芥、甘薯等植物中發現的基因,在植物抗寒性研究中都起到關鍵性的作用。
CBF/DREB1蛋白是一種廣泛存在于植物體內的控制植物體內低溫信號傳導的一類轉錄因子基因,也是現在研究最為廣泛的基因之一。近年來,隨著基因工程學科的不斷發展,從核桃中成功克隆出了許多影響核桃抗寒性的功能基因,相關研究只在研究單一低溫核桃相關基因的表達,但CBF基因在不同溫度處理表達的研究未見報道,因此,該課題對前人的研究進行了深入和完善,這對研究核桃抗寒機制是十分必要的。為了了解核桃內CBF基因對不同寒冷溫度的響應的變化,采用不同低溫梯度進行處理,利用RT-PCR技術研究了在不同時間不同溫度下下‘陽光’‘云新’核桃葉片內CBF基因在寒冷脅迫下表達量的變化規律,為以后選育核桃優良抗寒品種等提供更多的參考依據。
2 材料與方法
2.1 試驗材料
2.1.1 試驗材料
試驗于2015年12月~2017年3月在云南農業大學園林園藝學院果樹生理實驗室進行。試驗材料為云南農業大學園藝植物育種實驗室盆栽‘陽光’、‘云新’核桃幼苗葉梢對生葉片,樹齡0.5 a,處于幼年期,樹高1 m,沒有嚴重病害,在正常生長條件下‘陽光’長勢優于‘云新’核桃,但‘陽光’核桃更易受自然低溫和病害影響。‘云新’為云南薄殼與新疆核桃早實豐產雜交品種,‘陽光’為云南省選育的新品種。其中無病害室溫盆栽核桃作為空白對照,將整株核桃幼苗放入恒溫冰箱內進行活體2、4、6、8℃低溫脅迫處理,處理時間為2、4、8、24 h。處理完成后采摘新鮮葉片用錫紙包裹放入液氮浸泡,后放入超低溫冰箱保存。另一部分生理實驗材料,對處理后的新鮮核桃葉片立刻進行殺青、曬干、研磨成粉狀放入塑封袋密封保存。
2.1.2 試驗試劑
植物RNA提取試劑盒;cDNA第一鏈合成試劑盒、10×Loading Buffer、Marker為DL2000;2×Power Taq PCR MasterMix;氯仿、瓊脂糖等為國產分析純試劑。
2.1.3 儀器
移液器、大號研缽、冷凍離心機、水平電泳儀、凝膠成像系統,eppendorf熒光定量系統;將清潔后的研缽等用錫紙包裹后,置于200℃以上烤箱烘烤處理;96孔0.2 mL PCR管盒、巴洛克無RNA酶槍頭、八連管、1.5 mL離心管、1.5 mL離心管盒、0.2 mL PCR管、96孔冷凍鋁板。
2.1.4 PCR 引物設計與合成
進入NCBI網站,在GenBank數據庫中輸入物種拉丁名Juglans regia L.及相關基因名稱CBF獲得取該基因的DNA或mRNA序列后,以其為模板使用Primer Premier 5.0進行引物設計。引物設計完成后,委托昆明碩擎生物科技有限公司合成。
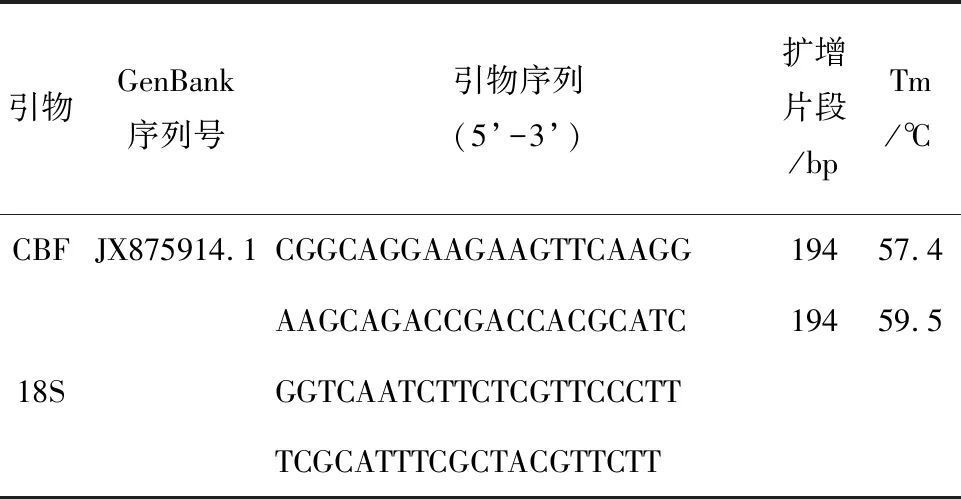
表1 qRT-PCR引物信息
2.2 方法
2.2.1 核桃葉片總 RNA 的提取
液氮覆蓋核桃新鮮葉片將其研磨成細粉末,待液氮揮發完后,用離心管估算裝取0.1 g樣品(可使用離心管稱取),加入1 mL細胞裂解液,在振蕩器上振蕩30秒后使其混勻后取1 mL勻漿物轉移至干凈的1.5 mL離心管中;在離心管加入300 μL去蛋白液和200 μL氯仿,在振蕩器上振蕩30 秒左右混勻;在常溫12 000 轉/分離心8分鐘后,將上清液轉移入干凈的1.5 mL離心管中,加入等體積的漂洗液,充分顛倒混勻,將混合物分2次在12 000 r/min常溫條件下離心1分鐘,棄去穿透液;加入500 μL洗柱液,12 000 r/min常溫離心1 分鐘,棄穿透液,重復一次該操作,然后常溫12 000轉/分離心1分鐘;將離心吸附柱轉移到干凈的1.5 mL離心管中,加入50 μL RNA洗脫液,放置片刻,12 000 r/min常溫離心1分鐘,離心管中溶液即為RNA樣品。分裝成5份,每份10 μL存放于超低溫冰箱待用。
2.2.2 RNA樣品電泳分析
取5 μL RNA樣品與等量的Buffer混勻,在1.5%瓊脂糖凝膠,1×TAE為電泳緩沖液,電壓為80 V的條件下電泳25 min,在凝膠成像儀上觀察RNA條帶呈像情況、并采集照片數據。
2.2.3 合成 cDNA 第一鏈
第一條鏈cDNA的合成按照PrimeScript RT reagent Kit with gDNA Eraser (TaKaRa公司)試劑盒說明書進行。
2.2.4 基因特異引物的PCR
驗證以cDNA為模板,用表1中的引物進行PCR擴增。PCR反應設定合適的循環參數,反應結束后4℃保存;擴增產物進行凝膠電泳檢測。
2.2.5 實時熒光定量PCR
樣本的制備后,上機進行樣品檢測。
2.2.6 數據處理
所有試驗重復3次,利用軟件DPS 7.05對組間進行單因素方差分析,利用Duncan’s 新復極差測驗。
3 結果與分析
3.1 核桃抗寒相關基因在‘陽光’‘ 云新’核桃葉片受到不同溫度和時間脅迫后的表達分析
3.1.1 總RNA質量及cDNA質量檢測
由圖1和圖3可見,RNA 28S和18S條帶亮度的比例約為2∶1,泳道無明顯背景表明RNA樣品的完整性較好、純度較高,無明顯降解,可用來接下來的的反轉錄。

1:2℃2H;2:2℃4H;3:2℃8H;4:2℃24H;5:4℃2H;6:4℃4H;7:4℃8H圖1 ‘陽光’核桃總RNA提取凝膠電泳圖像

1:6℃2H;2:6℃4H;3:6℃8H;4:6℃24H;5:8℃2H;6:8℃4H;7:8℃8H圖3 ‘陽光’核桃總RNA提取凝膠電泳圖像
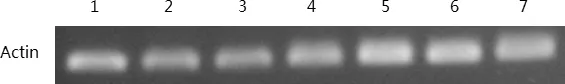
1:2℃2H;2:2℃4H;3:2℃8H;4:2℃24H;5:4℃2H;6:4℃4H;7:4℃8H圖4 ‘陽光’核桃樣品cDNA質量檢測凝膠呈像
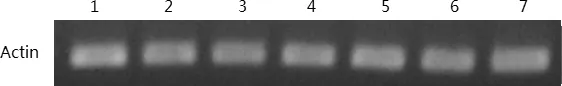
1:4℃24H;2:6℃2H;3:6℃4H;4:6℃8H;5:6℃24H;6:8℃2H;7:8℃4H圖5 ‘陽光’核桃樣品cDNA質量檢測凝膠呈像

1:8℃8H;2:8℃24H;3:常溫1號;4:常溫2號;5:常溫3號圖6 ‘陽光’核桃樣品cDNA質量檢測凝膠呈像

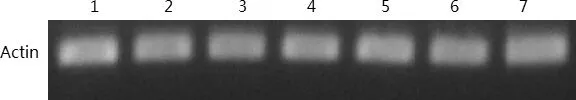
1:2℃2H;2:2℃4H;3:2℃8H;4:2℃24H;5:4℃2H;6:4℃4H;7:4℃8H圖7 ‘云新’核桃樣品cDNA質量檢測凝膠呈像
1:4℃24H;2:6℃2H;3:6℃4H;4:6℃8H;5:6℃24H;6:8℃2H;7:8℃4H圖8 ‘云新’核桃樣品cDNA質量檢測凝膠呈像
1:8℃8H;2:8℃24H;3:常溫1號;4:常溫2號;5:常溫3號圖9 ‘云新’核桃樣品cDNA質量檢測凝膠呈像
如圖7和圖9所示,以加入上述提取的‘云新’核桃葉片cDNA為模板,可擴增獲得1條清晰的片段,說明此方法提取的總RNA能夠滿足實時熒光定量PCR等以RNA為基礎的分子生物學實驗。
3.1.2 CBF基因在‘陽光’‘云新’核桃葉片受不同溫度脅迫后隨時間變化在葉片內的表達量分析
通過對‘陽光’核桃葉片組織進行實時熒光定量PCR檢測,發現CBF在核桃葉片中有表達,在三個不同時間CBF在核桃葉片內表達量均無明顯變化(圖10)。以核桃 18S 基因為內參,利用實時熒光定量PCR方法分析‘陽光’核桃葉片CBF在2、4、6、8℃四個低溫脅迫下隨時間延長核桃葉內基因表達量變化情況。其中在2℃條件下‘陽光’核桃葉片內的CBF基因表達量在隨著處理時間延長有較小幅度的升高,并在2 h處理時達到最高值,在4 h時有小幅下降,在8 h時又有小幅升高,處理24 h后基本不再變化(圖11)。4℃條件下‘陽光’核桃葉片內的CBF基因表達量在隨著處理時間延長有小幅變化,且變化并不明顯(圖12)。6 ℃條件下‘陽光’核桃葉片內的CBF基因表達量在隨著處理時間延長在2 h升高達到最高值后在4 h降低,處理8 h后又有小幅升高,24 h后又出現小幅降低(圖3-13),其中2 h處理與0、4、24 h處理相比差異性顯著。8 ℃條件下‘陽光’核桃葉片內的CBF基因表達量在隨著處理時間延長先升高,后隨著時間延長逐步降低,并在2H處理時達到最高值(圖14)與8 h、24 h處理相比差異性顯著。
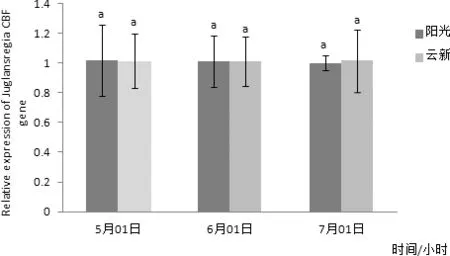
圖10 CBF基因在常溫下不同時間在兩個品種核桃葉片內表達量差異

圖11 CBF基因在2℃處理不同時間在兩個品種核桃葉片內表達量差異
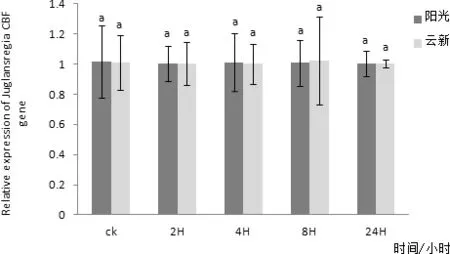
圖12 CBF基因在4℃處理不同時間在兩個品種核桃葉片內表達量差異
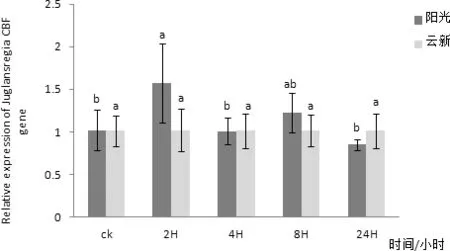
圖13 CBF基因在6℃處理不同時間在兩個品種核桃葉片內表達量差異
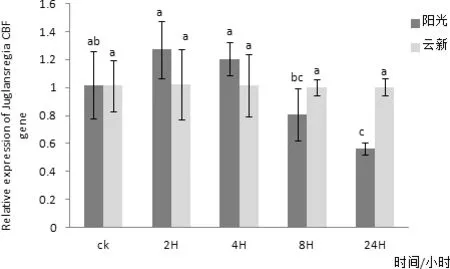
圖14 CBF基因在8℃處理不同時間在兩個品種核桃葉片內表達量差異
通過對‘云新’核桃葉片組織進行實時熒光定量PCR 檢測,發現CBF在核桃葉片中有表達,在三個不同時間CBF在核桃葉片內表達量均無明顯變化(圖3-11)。以核桃 18S基因為內參,利用實時熒光定量PCR方法分析‘云新’核桃葉片CBF在2、4、6、8 ℃四個低溫脅迫下隨時間延長核桃葉內基因表達量變化情況。但在2、4、6、8 ℃四個低溫條件下‘云新’核桃葉片內的CBF基因表達量在隨著處理時間延長均只有小幅變化,且變化不明顯(圖12~圖14)。在2℃低溫條件下‘云新’核桃葉片內的CBF基因表達量在隨著處理時間延長先降低后升高,然后趨于穩定。在4 ℃低溫條件下‘云新’核桃葉片內的CBF基因表達量在隨著處理時間延長先降低后升高之后再次降低,并在8 h達到最高值。在6 ℃低溫條件下‘云新’核桃葉片內的CBF基因表達量出現很小幅變化,在2 h達到最高值。在8 ℃低溫條件下‘云新’核桃葉片內的CBF基因表達量在隨著處理時間延長先升高后降低,并在2 h時達到最高值。
由以上實驗數據顯示,CBF在‘陽光’核桃低溫脅迫中起作用,在‘云新’核桃低溫脅迫中有表達,但表達量變化并沒有陽光核桃明顯。
CBF抗寒途徑是在高等植物中廣泛存在的抗寒抗旱、抗鹽堿信號傳導通路,植物通過CBF轉錄因子,激活下游COR、LTI、KIN基因的表達來提高植物抗寒性。李勇鵬從香樟中低溫誘導分離出了4種類CBF基因找到了控制香樟抵御逆境脅迫的重要轉錄因CcCBFa、CcCBFb、CcCBFc和CcCBFd[23]。吳純倩將擬南芥CBF3和CR15a冷誘導基因轉入煙草,提高了煙草的抗寒性及在低溫時的光合作用率[24]。雷恒久等通過平歐雜交榛克隆出ChaCBF1基因,通過研究發現該基因會使植株積累較高水平的可溶性糖和游離脯氨酸來提高自身抗寒能力[25]。馮勛偉等將從結縷草中克隆出了CBF同源基因ZjCBF轉入擬南芥中,發現即使不進行冷馴化ZjCBF過量表達的植株也比野生型抗寒性強[26]。張建朋等從麻核桃中克隆出jhCBF,序列分析說明JhCBF與樺木科白樺的親緣關系最近,優先聚為一類[27]。近些年來,研究者們從茶樹、巨桉(Eucalyptusgrandis)、葡萄、番茄(LycopersiconesculentumMill.)、扁桃(AmygdaluscommunisLinn.)等中都克隆出了類CBF基因,這些成果對研究抗低溫品種改良優化起著重要作用[28-32]。雷恒久[25]等對平歐雜交榛葉片進行4℃低溫處理發現ChaCBF1表達量隨時間延長均有升高。徐麗[33]等對‘香玲’核桃葉片在4℃脅迫后發現JrCBF基因隨處理時間延長出現了先升高后降低的趨勢。這些研究均說明CBF基因在植物抗寒過程中起到了重要作用。
‘陽光’核桃中CBF基因在6℃和8℃低溫脅迫下表達量有所升高,這暗示CBF基因在低溫脅迫過程中發揮重要作用。但‘陽光’核桃中CBF基因在4℃脅迫下并無明顯的表達量變化,而是在6℃和8℃時出現明顯變化,但受冷脅迫刺激后基因表達量變化趨勢相同。徐麗等所處的地區為山東省地理氣候環境與云南省地理氣候環境差異較大,是否能夠說明不同地域的不同品種的核桃的抗寒能力存在不同,所以導致CBF基因受低溫刺激表達變化的溫度區間不同。本實驗就云南省廣泛種植的‘云新’‘陽光’兩種核桃中CBF基因進行了低溫脅迫下表達量的研究。在實驗最后討論時發現‘云新’核桃在2℃~8℃CBF基因變化量不明顯,現階段并不能對討論所做的假設下定論。希望進一步實驗能夠對兩種核桃的綜合性抗寒指標進行測定,并選用不同樹齡植株材料,提供更大量的數據參考及使用更多品種核桃進行冷脅迫基因表達量分析,來推斷這種特殊情況是否只在‘云新’這個品種出現。
4 結論
結論:‘陽光’‘云新’核桃CBF基因在葉片不同溫度處理下均有表達,但各個時間段表達量之間有一定的差異。其中‘云新’核桃在2 、4、6、8 ℃四個處理溫度中表達量較為平均,沒有明顯變化,而‘陽光’核桃在6 ℃處理組中2 h處理后和8 ℃處理組中2 h處理后CBF基因的表達量較高。各個核桃產地均處于獨特的地理氣候環境中,也都有適應本土條件的核桃良種,核桃分子領域的研究基本為空白,希望通過這些研究為研究本土良種核桃抗寒生理及選育優良抗性核桃品種提供一定參考與思路。
猜你喜歡
中學生天地(A版)(2022年9期)2022-10-31 06:36:28
好日子(2022年3期)2022-06-01 15:58:27
故事作文·高年級(2021年10期)2021-10-23 13:21:26
動漫界·幼教365(中班)(2020年7期)2020-07-14 03:07:21
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
涼山文學(2016年6期)2016-12-05 11:51:42
少兒科學周刊·兒童版(2015年10期)2015-11-07 03:42:03
