基于MRI的早期盲人海馬體積分析研究
2019-05-23 06:55:46梁黎蓋新亭徐凱
放射學實踐 2019年5期
梁黎,蓋新亭,徐凱
視覺能夠使我們觀察周圍的環境、定位周圍的物體,并在其環境中導航。相關研究表明海馬涉及空間記憶及導航[1-2];也有研究表明海馬依賴于視覺及空間特征,來創建地圖式的空間模型,這些空間模型是導航的基礎[2]。在人類受試者中,通過磁共振功能成像研究發現海馬前部及右側海馬后部參與導航[3],且在導航任務期間海馬被激活[4]。由于缺乏正常人所容易獲得的視覺信息,盲人必須尋找一種替代方法來映射和導航周圍空間,并使用剩余的感官定位對象;因此盲人的海馬可能會發生一些適應性的改變。海馬的可塑性已經在許多動物實驗和人類研究中得到證實[5-7]。本文基于MRI的體積分析,探討海馬結構在早期盲人(先天性盲或嬰幼兒時期內失明)中可能發生的結構變化。

圖1 男,22歲,運用ITK-SNAP3.6.0軟件勾畫其左側海馬。a) 在軸面T1WI上勾畫; b) 在矢狀面T1WI上勾畫; c) 在冠狀面T1WI上勾畫; d) 三維立體結構圖。
1.研究對象
招募22例早期盲人,其中男13例,女9例,平均年齡(22.2±1.7)歲,均為右利手。同時選取22例視力正常者進行對照,其中男10例,女12例,平均年齡(21.9±1.1)歲。早期盲人入組標準:先天性失明或嬰幼兒時期內完全失明者,年齡20~30歲,右利手,所有受試者均有醫院相關證明文件。正常對照者入組標準:年齡與早期盲人組相匹配,自愿作為受試對象,具有完全正常的視功能(雙側裸眼視力大于1.0,具有雙側正常的視野及光敏度,具有正常的色覺)。所有受試者均排除糖尿病、心臟病、高血壓、精神病、遺傳病、頭部外傷史,無大量飲酒史,神經精神檢查無陽性發現,MRI圖像上無顱內病變表現。本研究方案獲本院倫理委員會批準,所有志愿者均簽署知情同意書。
2.研究方法
圖像采集:采用GE 3.0T磁共振掃描儀行MRI檢查,標準頭部線圈固定大腦,掃描野包括整個大腦,獲得3D-T1WI圖像。掃描參數:視野256 mm×256 mm,層厚1 mm,TR 1900 ms,TE 3.8 ms,矩陣256×256,體素1 mm×1 mm×1 mm,無間隔掃描176層。
圖像及數據處理:應用ITK-SNAP3.6.0軟件手動勾畫出海馬輪廓(圖1),并采用MATLAB軟件測量其體積。海馬定界方案參考Watson和Maller的定界方法[8-10],具體勾畫方法:由于海馬中灰質占優勢,主要勾畫顳角內側的T1WI等灰質信號區。海馬通常分為頭、體、尾三部分,海馬體部及尾部由于鄰近白質及腦脊液的襯托相對容易區分;但海馬頭部與鄰近結構分界欠清,在勾畫海馬頭部時以鉤狀隱窩作為海馬頭前上界標識,與杏仁體區分;同時下界與下托緊鄰的白質結構極易區分下托和海馬旁回;最后,外界為顳角內借下界向上的延伸區分海馬結構和鉤。為了減少誤差,每例每側海馬均由3位經過訓練的影像科醫師勾畫,運用MATLAB平臺測量體積后,取其平均值。組間比較時,為消除個體頭顱大小對海馬測量結果的影響,所有體積均進行標準化處理,體積標準化值=(海馬體積/個體顱腔體積)×顱腔體積平均值,顱腔體積平均值為本研究所有受試者的顱腔體積的平均值。參考相關文獻[11,12],將海馬前1/4劃為前部,后1/4劃為后部,剩下為中部。
3.統計學分析
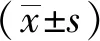
結 果
1.組內海馬總體積比較
正常對照組及盲人組左、右側海馬總體積均符合正態分布。正常對照組及盲人組左、右側海馬總體積的比較采用配對樣本t檢驗,結果顯示正常對照組、盲人組右側海馬總體積均明顯大于左側,差異均有統計學意義(P<0.05,表1)。
2.組間海馬總體積及各部分體積比較
正常對照組、盲人組左、右兩側海馬總體積及各部分體積的比較采用獨立樣本t檢驗,結果顯示正常對照組左、右側海馬總體積均大于早期盲人組,差異均有統計學意義(P值均<0.05,表2)。早期盲人組右側后部及左側中部體積小于正常對照組,差異有統計學意義(P值<0.05,表3、圖2)。

圖2 正常對照組與早期盲人組間海馬各部分體積比較。標準差用柱狀圖上垂直線條表示,“*”表示差異有統計學意義(P<0.05)。

表1 組內海馬總體積比較(mm3)

表2 標準化后組間海馬總體積比較(mm3)

表3 標準化后組間各部分海馬總體積比較(mm3)
討 論
海馬體積的研究是評價海馬正常生理狀態和病理變化的基礎,對相關臨床和科學研究有重要意義[9,13]。雖然海馬結構在多種疾病中被廣泛研究,但在失明患者中的相關研究較少。Chen等[14]在使用基于體素的形態學研究中發現,雙側晚期青光眼患者中海馬回灰質體積顯著降低。Chebat等[11]研究發現盲人右側海馬后部縮小,Fortin等[15]報道盲人右側海馬頭增大。Maller等[16]發現與正常對照組相比,盲人海馬總體積及區域海馬體積無明顯差異。這些研究可能反映記憶系統的功能需求增加和/或對視覺剝奪的適應性反應。
本研究基于3.0T MRI掃描獲得3D-T1WI圖像,約50%以前的結構研究使用了低場強MRI掃描儀(1.5T)。3.0T MRI具有空間及時間分辨率高、信噪比高、偽影小等優點[17]。同時采用手動分割方法,目前海馬的手動分割被視為所有分割方法的金標準[7],相對于自動分割的低精確度,手動分割仍具有優勢[18]。
本研究比較了正常對照組及早期盲人組左、右側海馬總體積,發現兩組的右側海馬總體積均明顯大于左側,差異有統計學意義,可能與研究對象均為右利手有關[8]。提示無論在正常對照組還是早期盲人組中右側海馬體積仍保持優勢半球功能。
本研究也對早期盲人與正常人兩側海馬總體積進行了組間比較,結果顯示早期盲人組左、右側海馬總體積均低于正常對照組,差異均有統計學意義;這可能是早期盲人視覺剝奪后的適應性反應。筆者進一步分析了早期盲人組與正常對照組的兩側前部、中部及后部體積,發現早期盲人組右側海馬后部及左側海馬中部體積較正常對照組小,差異有統計學意義。這與Szabo等[10]的研究結果部分一致,而與Fortin等[15]、Maller等[16]的研究結果相反,可能與他們采用低場強MRI(1.5T)、樣本量較小有關。海馬中間區域及后部與空間導航有關[19-20],而視覺信息對于空間導航至關重要[2]。海馬中間區域通過內嗅皮層的背外側和中間帶與對精準空間導航很重要的視覺及軀體感覺皮層連接[19]。相關學者在導航任務研究中發現海馬后部涉及視覺空間編碼和路徑中物體位置的檢索[4]。因此,視覺剝奪可能直接導致盲人海馬中部和后部與視覺空間導航有關的神經元發生廢用性萎縮,這可以解釋早期盲人左側海馬中部、右側海馬后部體積縮小。
盡管有研究強烈暗示海馬參與視力正常人的空間導航[21-22],但對于盲人而言,盲人不一定在導航能力方面顯示出障礙。相關研究發現相對于正常人,盲人表現出相當或甚至優越的非視覺空間導航能力[15,23]。Passini等[24]研究發現,盲人仍能導航并形成認知空間,表明海馬在空間導航中與其他大腦區域共享。也有研究表明情景記憶以及空間導航依賴于皮質-海馬的相互作用[19,25]。Kravitz等[26]也發現在空間導航過程中,海馬很少單獨被激活,通常與其他大腦區域一起運行,例如前額葉和頂葉皮質。Ma等[27]研究表明,先天性盲人海馬與頂枕溝、眶額葉前腦皮質和額中回靜息態功能連接增加。而且,頂葉皮層參與空間處理及空間記憶,內側頂葉可能在及時空間運動中發揮重要作用[28]。因此,盡管右側海馬后部及左側海馬中部體積變小,早期盲人仍可通過其他大腦區域和/或功能連接的增強來進行導航,提示在早期盲人空間導航中較少依賴海馬。
綜上所述,本研究結果表明早期盲人組及正常對照組的右側海馬總體積均大于左側,且早期盲人組的兩側海馬總體積均低于正常對照組,以右側海馬后部和左側海馬中部為著;表明早期盲人由于視覺剝奪后兩側海馬可能發生適應性改變,但仍然保持右側優勢半球功能。本研究結果可為將來研究盲人空間導航或認知功能變化提供相關的結構基礎,并進一步為臨床診療提供參考依據。
猜你喜歡
體育科技文獻通報(2022年3期)2022-05-23 13:46:54
作文周刊·小學二年級版(2022年20期)2022-05-05 01:33:06
天津外國語大學學報(2021年3期)2021-08-13 08:32:18
遼金歷史與考古(2021年0期)2021-07-29 01:06:54
科技傳播(2019年22期)2020-01-14 03:06:54
創新作文(小學版)(2019年10期)2019-09-25 08:12:28
民用飛機設計與研究(2019年4期)2019-05-21 07:21:24
汽車工程學報(2017年2期)2017-07-05 08:13:02
小學生學習指導(低年級)(2017年5期)2017-05-04 04:14:38
大灰狼(2015年6期)2015-07-16 21:01:00
