海燕視覺的初步研究
2019-05-28 00:26:59張繼文田濤姜城方星楠孫宇范繼涵曲鵬健
安徽農學通報 2019年8期
張繼文 田濤 姜城 方星楠 孫宇 范繼涵 曲鵬健


摘? 要:海燕(Asteroidea)是棘皮動物門海星綱有棘目海燕科的一種海洋類無脊椎底棲動物,主要分布于我國渤海和黃海一帶。海燕喜食軟體動物,包括一些貝類的軟體組織,已成為貝類和貝類養殖的敵害。通過實驗方法研究海燕的視覺行為,可以掌握其行為規律,為采取有效措施,減少海燕對貝類養殖的危害提供科學依據。使用黑、紅、藍、綠4種顏色的水槽對海燕進行了視覺實驗分析,結果表明,海燕對黑、紅、藍、綠4種顏色具有一定的選擇性,在正常顏色選擇實驗中,海燕在不同顏色停留時間為黑色>藍色>紅色>綠色。使用4種顏色對海燕進行攝食馴化,紅色和藍色隨著攝食馴化的時間增長,海燕在這2種顏色停留的時間越長,黑色和綠色的馴化結果并不理想,隨著馴化時間的增長,海燕在這2種顏色區域停留的時間并沒有加長。實驗證明,海燕對顏色具有一定的選擇性,具備一定的視覺能力。
關鍵詞:海燕;視覺;攝食馴化;顏色選擇
中圖分類號 Q959.7 文獻標識碼 A 文章編號 1007-7731(2019)08-0048-05
Abstract:Asteroidea is a phylum of echinoderms,Neptunidae,Acanthoideae,a marine invertebrate benthic animal.It mainly distributes in the Bohai Sea and the Yellow Sea of China.The Asteroidea likes mollusks,including some mollusks,therefore,the petrel has become an enemy of shellfish and shellfish culture.Therefore,Asteroidea has become a predator of shellfish and shellfish farming.The experimental study of Asteroidea 's visual behavior can further prevent the damage of Asteroidea to culture,povide a scientific basis for measures taken to reduce the hazards to cultured shellfish.Visual experiment analysis of Asteroidea using tank of four color:black,red,blue and green.The results show that the Asteroidea has a certain selectivity to the four colors of black,red,blue and green.In the normal color selection experiment,the residence time of Asteroidea in different colors was black>Blue>Red>green.Feeding and domesticating Asteroidea in four colors,Red and blue grow with the time of food and domestication,and the longer the Asteroidea stays in these two colors,The domestication results of black and green are not ideal.As the domestication time increases,Asteroidea's stay in these two color areas does not lengthen.The experiment proves that Asteroidea has certain selectivity to color,and Asteroidea has certain visual ability.
Key words:Asteroidea;Vision;Feeding and domestication
海燕(Asteroidea)是棘皮動物門海星綱有棘目海燕科[1]的一種海洋類無脊椎底棲動物,生活在潮間帶及沿海的淺水帶,主要分布于我國渤海和黃海一帶[2]。多生活于沿岸淺海或潮間帶巖石海岸,常裸露,或隱藏在石下或石縫中,有少數種潛伏在沙灘表面。屬于掠食性動物,食量較大,主要攝食軟體動物,喜食菲律賓蛤仔、扇貝、貽貝、鮑魚等,因此海燕成為了貝類和貝類養殖的敵害[3],特別是對雙殼貝類具有很大的威脅性,對經濟貝類的養殖以及潮間帶生物多樣性都有著重大影響[4-7]。
對于棘皮動物的視覺研究國外很早就開始進行了,尼爾森將動物的視覺描述為以下4個進化階段:(1)簡單的光源接收;(2)具有一定方向性的光接收,允許基本的趨光性;(3)低分辨率空間視覺;(4)高分辨率多用途視覺[8]。目前只在3種動物中的視覺在能力達到了第4個階段,分別是頭足類軟體動物,節肢動物和脊椎動物[9-11]。已經被證實棘冠海星存在能夠形成視覺的復眼,并且復眼能夠起到導航作用,刺冠海星具有迄今為止所研究的任何海星中最高的空間分辨率。此外,它們是檢查的所有動物視力最慢的,閃爍融合頻率僅為0.6~0.7Hz。這可以是自適應的,因為檢測諸如珊瑚礁的靜止物體不需要快速視覺[11]。研究海燕的視覺能夠揭示其對海洋生物影響以及其行為的作用基礎。為此,本研究以海燕為實驗對象,研究其對不同顏色的選擇,并通過攝食馴化來觀察海燕是否對顏色有識別能力,了解其視覺能力,為進一步控制海燕對養殖經濟貝類的危害和海星的誘捕剔除提供科學依據。
1 材料與方法
1.1 實驗材料 實驗所用海燕為從遼寧省大連市獐子島海域使用地籠網捕獲的,餌料菲律賓蛤仔取自遼寧省大連市黑石礁海域。圓形水槽(內徑0.9m,高0.4m)購買自大連塑料城,塑料防水彩紙為網購,共有黑、紅、藍、綠4種顏色。使用高清攝像機進行實驗錄像拍攝。使用塑料防水貼紙將圓形水槽按照4種顏色各90°扇形將水槽貼成4種顏色,并在中心位置留出一塊直徑為0.2m圓形空白區域(圖1)。
1.3 實驗設計
1.3.1 顏色選擇實驗 將四色圓形水槽放置于攝像機下,每天早上9:00隨機選取3只海燕,將海燕背面朝上放置于水槽中的白色區域內,進行24h錄像。第2天將3只海燕取出,更換海水,并將水槽順時針轉動90°,隨機選取3只海燕,將海燕背面朝上放置于水槽中的白色區域,進行24h錄像,實驗進行4d。實驗第5天,改變水槽中顏色排列的順序,同樣每天選取3只海燕進行實驗,錄像24h,實驗再進行4d,每天都順時針90°改變水槽的顏色位置。
1.3.2 顏色馴化實驗 實驗開始時,隨機選擇3只海燕背面朝上放置于水槽的白色區域中,每天早上7∶00,在紅色區域內投喂菲律賓蛤仔,每天下午7∶00將水槽中的菲律賓蛤仔取出并換水,投喂新的菲律賓蛤仔。馴化1d后,將3只海燕取出,背面朝上放置于錄像機下的水槽中,進行24h的實驗錄像。分別在馴化3d、5d、7d后進行24h的實驗錄像。接下來分別在綠色、藍色、黑色區域內進行海燕的馴化,分別在馴化1、3、5、7d后進行24h的實驗錄像。
1.4 數據處理及分析 試驗數據均利用SPSS 17.0軟件對測定結果進行統計分析,實驗數據用平均值±標準誤差(Mean+SE),采用方差分析和Duncan氏檢驗進行顯著性分析和多重比較,數據在P<0.05時被認為具有顯著性差異,反之則無差異性。
2 結果與分析
2.1 海燕對不同顏色的選擇 實驗進行了8d,海燕在不同顏色區域停留的時間如圖2所示。從圖2可以看出,海燕對顏色的選擇趨向為黑色>藍色>紅色>綠色(白色部分為實驗開始放置區域,不計入顏色選擇)。從圖3可以看出,海燕在不同區域中的停留時間最高的黑色占27.2%,最少的綠色占14.9%(因為白色為海燕放置區域,并且顏色面積與其他4種顏色的大小不同,所以不將白色算在顏色趨向內)。
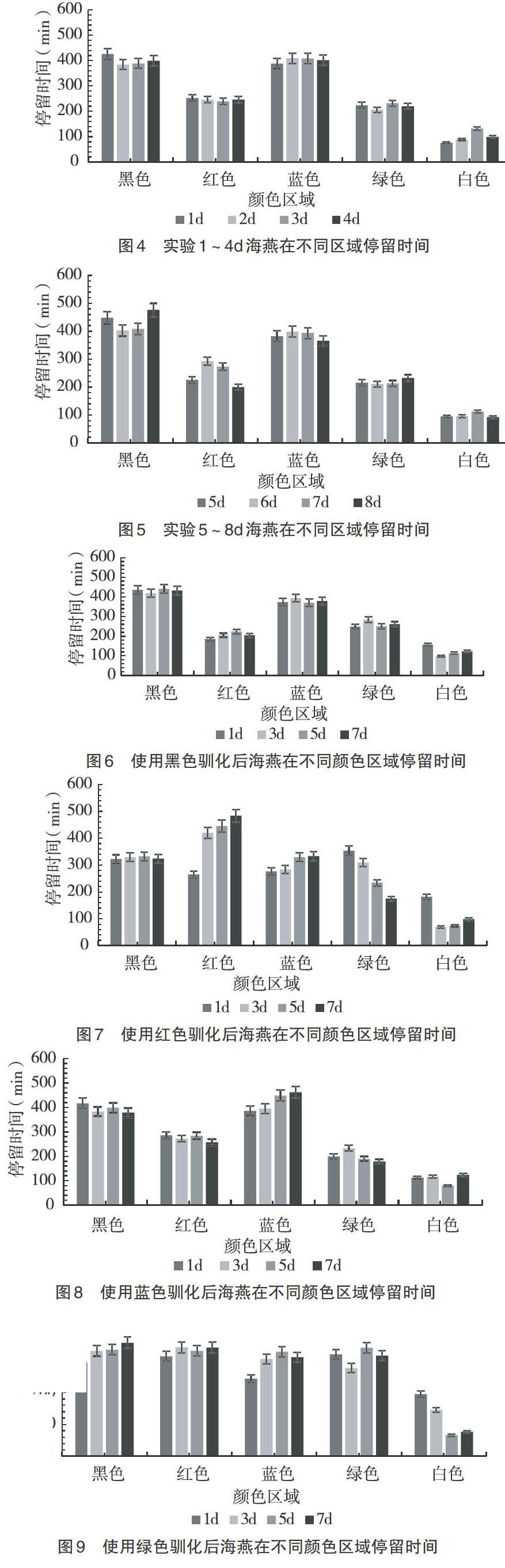
在水槽顏色未改變顏色的1~4d實驗和水槽改變顏色5~8d實驗中,海燕在不同顏色區域停留的時間如圖4、5所示。從圖4、5可以看出,無論是1~4d實驗還是5~8d實驗,隨著水槽的90°轉動,海燕在不同的顏色區域停留的時間會發生一定的波動,但是并沒有在很大程度上影響海燕對于這4種顏色選擇的趨向,海燕對于顏色的趨向仍是黑色>藍色>紅色>綠色。在改變水槽的顏色排列順序之后,海燕在不同顏色區域停留的時間也會產生一定的波動,但是海燕對于4種顏色的趨向并未發生改變,可以排除實驗中地磁以及光源方向對實驗產生的影響。
2.2 馴化后的海燕對顏色的選擇
2.2.1 黑色馴化 使用黑色對海燕進行攝食馴化1、3、5、7d后,海燕在不同顏色區域停留時間的情況如圖6所示。從圖6可以看出,隨著使用黑色區域對海燕進行攝食馴化1、3、5、7d后,海燕在不同顏色的區域的停留時間會發生一定的波動,但是整體上來說并沒有改變海燕對顏色的趨向關系,同時海燕在黑色區域的停留時間也沒有出現明顯的上升或者下降趨勢。
2.2.2 紅色馴化 使用紅色對海燕進行攝食馴化1、3、5、7d后,海燕在不同顏色區域停留時間的情況如圖7所示。從圖7可以看出,在馴化1d的時候,海燕會出現顏色選擇紊亂的現象,原本的顏色趨向被打破,綠色區域的停留時間明顯上升。隨著馴化時間的加長,可以看到,海燕在紅色區域停留的時間也明顯加長,海燕在紅色區域攝食馴化之后,隨著馴化時間的增加,海燕在紅色區域停留的時間增加。
2.2.3 藍色馴化 使用藍色對海燕進行攝食馴化1、3、5、7d后,海燕在不同顏色區域停留時間的情況如圖8所示。從圖8可以看出,使用藍色對海燕進行攝食馴化,第1天時,并沒有表現海燕原本對于不同顏色的趨向,也沒有出現顏色選擇紊亂的現象。隨著馴化時間的增加,海燕在藍色區域停留的時間逐漸增加,使用藍色馴化第3天的時候,海燕在藍色區域停留的時間超過海燕在黑色區域停留的時間,最后海燕對于不同顏色的停留時間變成藍色>黑色>紅色>綠色。
2.2.4 綠色馴化 使用綠色對海燕進行攝食馴化1、3、5、7d后,海燕在不同顏色區域停留時間的情況如圖9所示。從圖9可以看出,使用綠色對海燕進行攝食馴化1、3、5、7d,海燕一直出現顏色選擇紊亂的情況,隨著馴化時間的增加,海燕在不同顏色區域停留的時間沒有出現明顯的趨勢,只有在黑色區域停留的時間隨著馴化時間的增加而增大。
2.2.5 顏色馴化效果 海燕對于4種顏色區域的攝食馴化隨著馴化時間增長,在不同馴化顏色區域停留的時間變化如圖10所示。從圖10可以看出,隨著馴化時間的增加,海燕紅色和藍色區域停留的時間是呈上升趨勢的,而且紅色的上升趨勢更加明顯,上升幅度也很大。海燕在黑色區域停留的時間整體上都很高,但是隨著時間的增加并沒有呈現上升的趨勢,海燕在綠色區域的停留時間波動很大,也沒有呈現上升或者下降的趨勢。
3 討論
3.1 棘皮動物的視覺 一直以來,棘皮動物都被認為是不存在視覺的,大部分研究學者都認為棘皮動物主要靠嗅覺取定位和搜尋食物。但是,Garm和Nilsson已研究表明,在海星Linckia laevigata中,眼睛被用來定位大型地標,特別是它們通常居住的珊瑚礁的黑暗邊緣[12]。同時Nilsson將動物的視覺描述為產生于以下4個進化階段:第1階段,簡單的光接收;第2階段,具有一定方向性的光接收,允許基本的趨光性;第3階段,低分辨率空間視覺;第四階段,存在高分辨率多用途視覺[8]。在5類棘皮動物中均發現了皮下光感覺接受,但是真正的存在能夠具有成像的視覺的眼睛,只在1種海參和幾種海星中發現。海燕存在一定的光接收能力,并且它的趨暗性也表現得很明顯。很多國內文獻對海燕攝食的晝夜節律研究表明,海燕在夜間的攝食能力明顯高于白天[13-19]。本實驗中,可以明顯發現,海燕對于黑色和藍色區域的趨向性,同時在不同顏色區域停留的時間的長短也能表現海燕對不同顏色存在一定的選擇性。在馴化實驗中,海燕表現出對于紅色和藍色更加優秀的識別能力,通過攝食馴化,海燕能夠隨著馴化時間的加長,分辨出投喂食物的顏色區域,并且在改顏色區域停留的時間明顯加長,從中也能看出海鹽具有一定的視覺能力。
3.2 視覺位置 棘皮動物的視覺位置大部分都屬于皮下組織的光感受器,只有幾種棘皮動物存在真正的形成視覺的眼睛。棘冠海星存在能夠形成視覺的真正的“眼睛”,它的眼睛是和昆蟲一樣的復眼,復眼的位置在腕足前端的一個小凸起上,棘冠海星可以通過改變凸起的角度,來調整自己視角的方向。海燕并沒有存在明顯的復眼結構,它的腕足前段也存在一些觸角,但是很多學者認為這是海燕的嗅覺感受器,和視覺并不存在聯系,所以海燕也可能是依靠皮下組織的光感受器來進行光的接受和顏色的分辨的[20]。這也是很多棘皮動物的視覺存在形式,海燕的視覺在正常海底生活時用處不大,因為生存的環境,光纖并不充足,同時海燕不存在成像視覺的“眼睛”,大部分移動和攝食還是需要靠嗅覺來完成,這樣的情況也和棘冠海星類似,雖然它具有能夠成像的復眼,但是這個成像距離很近,只能在1~2m以內形成視覺,敷衍的駐顏作用還是用來尋找珊瑚礁的大概位置,遠一點的位置也是要靠嗅覺來完成定位[21-23]。
3.3 視覺功能和進化 Nilsson提出視覺的演變已經從非定向的光接收到具有高空間分辨率的功能進化時產生的,并且每個階段都能夠擁有越來越先進的視覺引導能力[24]。根據這個理論,棘皮動物的視覺能力都是存在的,都能夠進行一些簡單的視覺引導行為,而且只需要非常簡單的視覺系統處理[25]。同時,這也是棘皮動物的視覺進化的方向,通過不斷的進化,棘皮動物會游泳更先進的視覺能力。目前,在很多的動物身上都發現了非常發達的視覺能力,扇貝具有2°的視覺分辨率,alciopid和heteropod的視覺分辨率在1°左右,這樣看起來它們和一些昆蟲的視覺分辨率一樣好,但是像這幾種動物的視覺能力和已經具有發達視覺能力的頭足類軟體動物、節肢動物和脊椎動物相比并不是那么突出,對于視覺的引導行為也不那么明顯[26]。其原因主要是因為后者具有較大的大腦,而在大腦中元用于視覺的部分所占比例也很大。簡單來說,這些動物的大腦或者神經元并沒有具備能夠完全發揮其視覺的全部潛力,這也是棘皮動物的視覺進化的方向[27-30]。
3.4 海燕的行為 本實驗主要觀察了海燕對不同顏色的選擇以及是否存在視覺記憶,同時試驗過程中也觀察到了很多海燕的別的行為。比如,海燕存在“追逐”行為,海燕會在選定一個顏色后運動停止,這時當另外一只海燕靠近這只停止運動的海燕時,就會發生這種“追逐行為”,停止運動的海燕會在運動過來的海燕的追逐中重新開始運動。海燕在水槽壁上運動的過程中,偶爾還會存在掉落的現象,掉落后的海燕會隨機選擇顏色運動,并沒有一定的規律,這樣的現象可能是應激反應的一種表現,深入研究還需要更多的實驗進行觀察和總結。
4 結論
(1)海燕對黑、藍、紅、綠4種顏色具有一定的趨向性,在不同顏色區域的停留時間長短不同,顏色選擇趨向為黑色>藍色>紅色>綠色。地磁和燈光陰影并不會改變海燕對于這4種顏色的選擇情況。
(2)使用4種顏色對海燕進行攝食馴化,隨著攝食馴化的時間增長,海燕在紅色和藍色這2種顏色停留的時間越長;而黑色和綠色的馴化結果并不理想,隨著馴化時間的增長,海燕在這2種顏色區域停留的時間并沒有加長。
(3)從馴化效果來看,紅色的馴化效果比藍色更明顯。
參考文獻
[1]裴青.海燕化學成分的分離與鑒定[J].沈陽藥科大學學報,2012,29(10).
[2]郭承華.海星資源的開發利用[J].生物學通報,1989,11.
[3]郭文場,劉佳賀,趙冰冰.中國海星資源的種類、分布與綜合利用[J].特種經濟動植物,2013,12.
[4]Sloan N A,Aspects of the feeding biology of animals[J].Oceanography and Marine Biology Annual Review,1980,18:57-124.
[5]Power M E,Tilmao D,Estes B A,et al.Challenges in the quest for keystones[J].Bioscience,1996,46:609-620.
[6]Witman J D,Dayton P K.Rocky subtidal communities//Bertness,M.D.,Gaines,S.D.,Hay,M.E.eds.Marine Community Ecology[M].Sinauer Associates,Massachusetts,2001:339-366.
[7]Verling E,Crook A C,Barnes D K A,et al.Structural dynamics of a sea star Marthasterias glacialis population[J].Marin Biological Association of the United Kingdom,2003,83:583-592.
[8]Land,Michael F.Animal Vision:Starfish Can See at Last[J].Current Biology,2014,24(5):200-201.
[9]Garm A ,Nilsson D E .Visual navigation in starfish:first evidence for the use of vision and eyes in starfish[J].Proceedings of the Royal Society B:Biological Sciences,2014,81(1777):20133011-20133011.
[10]Nilsson D E .The evolution of eyes and visually guided behaviour[J].Philosophical Transactions of The Royal Society B Biological Sciences,2009,364(1531):2833-2847.
[11]Nilsson,D.-E.Eyes as Optical Alarm Systems in Fan Worms and Ark Clams[J].Philosophical Transactions of the Royal Society B:Biological Sciences,1994,346(1316):195-212.
[12] Land, M.F.,Nilsson, D.-E.General-purpose and special-purpose visual systems. In Invertebrate Vision, E. Warrant and? D.-E. Nilsson, eds[J].Cambridge:Cambridge University Press,2006:167-210.
[13]Himmelman J H,Dutil C,Gaymer C F.Foraging behavior and activity budgets of sea stars on a subtidal sediment bottom community[J].Journal of Experimental Marine Biology & Ecology,2005,322(2):153-165.
[14]Holling C S.The Functional Response of Invertebrate Predators to Prey Density1[J].Memoirs of the Entomological Society of Canada,1966,98(48):1-86.
[15]Rice J A,Crowder L B,Marschall E A.Predation on juvenile fishes:dynamic interactions between size-structured predators and prey[M]// Early Life History and Recruitment in Fish Populations.Springer Netherlands,1997.
[16]張天文,劉廣斌,劉恩孚,等.多棘海盤車對魁蚶攝食量、選擇性及晝夜攝食差異的初步研究[J].中國海洋大學學報(自然科學版),2015,45(12):24-29.
[17]代克濤,李嬌,關長濤,等.多棘海盤車(Asterias amurensis)對5種雙殼貝類的攝食選擇性及攝食節律[J].漁業科學進展,2015,36(01):97-102.
[18]劉佳.多棘海盤車攝食行為和繁殖生物學的初步研究[D].青島:中國海洋大學,2012.
[19]齊占會,王珺,毛玉澤,等.兩種海星對三種雙殼貝類的捕食選擇性和攝食率[J].生態學報,2013,33(16):4878-4884.
[20]Borst A .Current Biology-Comments on Drosophila's View on Insect Vision[J].Current Biology.
[21]Anders G,Jan B,Ronald P,et al.Hunting in Bioluminescent Light:Vision in the Nocturnal Box Jellyfish Copula sivickisi[J].Frontiers in Physiology,2016,7.
[22]Bos A R,Gumanao G S,Mueller B,et al.Management of crown-of-thorns sea star(Acanthaster planci L.)outbreaks:Removal success depends on reef topography and timing with in the reproduction cycle[J].Ocean & Coastal Management,2013,71(1):116-122.
[23]Riveraposada J,Pratchett M S,Aguilar C,et al.Bile salts and the single-shot lethal injection method for killing crown-of-thorns sea stars(Acanthaster planci)[J].Ocean & Coastal Management,2014,102:383-390.
[24]Nilsson DE.2004 Eye evolution:a question of genetic promiscurity.Curr.Opin.Neurobiol.14,407-414.(doi:10.1016/j.conb.2004.07.004)
[25]Arendt D,Wittbrodt J.Reconstructing the eyes of Urbilateria[J].Philosophical Transactions of the Royal Society of London,2001,356(1414):1545-1563.
[26]Penn P E,Alexander C G.Fine structure of the optic cusion in the asteroid Nepanthia belcheri[J].Marine Biology,1980,58(4):251-256.
[27]Ullrich-Luter E M,Dupont S,Arboleda E,et al.Unique system of photoreceptors in sea urchin tube feet[J].Proc Natl Acad Sci U S A,2011,108(20):8367-8372.
[28]Raible F,Tessmar-Raible K,Arboleda E,et al.Opsins and clusters of sensory G-protein-coupled receptors in the sea urchin genome[J].Developmental Biology,2006,300(1).
[29]Burke RD. A genomic view of the sea urchin nervous system[J].Dev.Biol,2006,300,434-460.
[30]Nilsson D.-E.The evolution of eyes and visually guided behaviour[J].Proc. R. Soc.B,2009,364,2833-2847.(doi:10.1098/rstb. 2009.0083) (責編:張宏民)
