
黃酒熟麥曲中細菌多樣性的評價
2019-05-31 05:19:36崔夢君折米娜張振東趙慧君郭壯
食品研究與開發 2019年11期
崔夢君,折米娜,張振東,趙慧君,郭壯
(湖北文理學院食品科學技術學院 鄂西北傳統發酵食品研究所,湖北 襄陽 441053)
作為黃酒釀造中的重要原料,麥曲中包含霉菌、酵母菌和細菌等微生物,由于微生物與生物環境的互生、共生、寄生和拮抗等關系,逐漸形成了一個復雜且具有多樣性的微生物區系,同時對黃酒風味也起到了至關重要的作用[1-2]。譚婷婷等從北方黃酒麥曲中分離出5 株霉菌和1 株酵母菌,分別為多枝橫梗霉、米曲霉、廣紫青霉、雜色曲霉、交鏈孢霉和釀酒酵母[3],曹玨等發現米根霉、微小毛霉、米曲霉和煙曲霉是紹興黃酒麥曲中的主要真菌[4]。目前關于黃酒發酵過程中揮發性風味物質的變化以及真菌群落結構進行了大量研究[5-7],然而關于麥曲中細菌多樣性的研究卻鮮有報道。
變性梯度凝膠電泳(denatured gradient gel electrophoresis,DGGE)是一種能夠快速對環境中的微生物群落進行評估的分子生物學技術[8],被廣泛應用于食醋[9]、泡菜[10]、香腸[11]、大醬[12]和黃酒[13]等發酵食品的微生物多樣性解析中。Illumina MiSeq 是一種高通量測序方法,可以以相對較低的成本產出大量序列,與其他二代高通量技術相比增加了微生物群落分析的測序深度,同時降低了試驗成本,使其成為根據大規模標記基因測序研究不同環境中微生物群落的新方法[14-15],目前在農業飼料[16]、發酵食品[17]和環境衛生[18]微生物解析領域均有廣泛的應用。
本研究以黃酒麥曲為研究對象,在提取樣品微生物宏基因組DNA 的基礎上,利用DGGE 與MiSeq 高通量測序技術相結合的手段對其細菌多樣性進行評價,同時結合生物信息學手段對其微生物群落結構進行解析,以期為后續麥曲中微生物資源的挖掘提供理論支持。
1 材料與方法
1.1 材料與儀器
熟麥曲:采集自浙江省麗水市,編號分別為MQ1、MQ2、MQ3 和 MQ4;三羥甲基氨基甲烷、乙酸、乙二胺四乙酸、丙烯酰胺、甲叉雙丙烯酰胺、去離子甲酰胺、尿素、過硫酸銨、四甲基乙二胺、乙醇、冰醋酸、甲醛、硝酸銀、氫氧化鈉:國藥集團化學試劑有限公司;D5625-01 DNA 提取試劑盒、DNA marker、聚合酶鏈式反應(polymerase chain reaction,PCR)清潔試劑盒:京科博匯智生物科技發展有限公司;2PCR×mix:南京諾唯贊生物科技有限公司;rTaq、2'-脫氧核苷酸-5'-三磷酸混合物(dNTP MIX)、pMD18-T vector:大連寶生物技術有限公司;引物:武漢天一輝遠生物科技有限公司。
引物信息見表1。
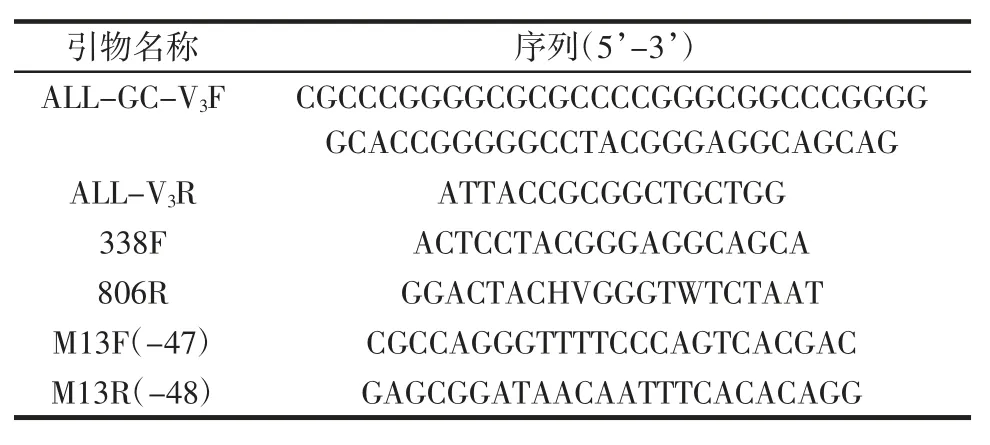
表1 引物信息Table 1 The information of primers
VeritiTM96-well Thermal Cycler PCR 儀:美國AB公司;NanoDrop 2000:美國 Thermo Fisher 公司;DCodeTMSystem:美國 Bio-Rad 公司;DYY-12 電泳儀:北京六一儀器廠;MiSeq PE300 高通量測序平臺:美國Illumina 公司;R920 機架式服務器:美國DELL 公司;CT15RE 冷凍離心機:日本HITACHI 公司;Bio-5000 plus 掃描儀:上海中晶科技有限公司。
1.2 方法
1.2.1 樣品微生物宏基因組提取與檢測
采用D5625-01 試劑盒提取麥曲微生物宏基因組DNA,用0.8%瓊脂糖凝膠進行電泳檢測并使用NanoDrop 測定濃度。
1.2.2 DGGE 電泳及優勢條帶測序
將樣品總DNA 作為模板進行聚合酶鏈式反應(PCR),采用細菌通用正向引物ALL-GC-V3F 和反向引物ALL-V3R 對樣品16S rDNA V3 區域進行擴增。PCR 擴增總體積為 25 μL,包括 10×PCR Buffer(含Mg2+)2.5 μL,dNTP 2 μL,正、反向引物各 0.5 μL,rTaq 0.5 μL,模板 1 μL,無菌超純水補至 25 μL。PCR 擴增程序:95℃預變性 4 min,95℃變性 30 s,55℃退火30 s,72℃延伸 30 s,循環 30 次,最后 72℃終延伸10 min。所得擴增產物用2%的瓊脂糖凝膠電泳檢測。使用Bio-Rad 公司Dcode 突變檢測系統對上述PCR產物進行電泳,采用8%的丙烯酰胺凝膠,變性梯度為35%~52%。DGGE 條件:恒溫 60℃,0.5 TAE 電泳緩沖液,上樣量 10 μL,電壓先 120 V,運行 80 min,后電壓80 V,運行780 min。電泳結束后,采用硝酸銀法染色,使用掃描儀將電泳圖進行拍照,找出各樣品特征條帶并回收膠塊。用不帶GC 夾板的引物(ALL-V3F 和ALL-V3R)將回收膠塊再次進行PCR 擴增。試劑盒純化PCR 產物并進行載體連接和克隆培養,挑取陽性克隆子送往武漢天一輝遠生物科技進行測序。
1.2.3 麥曲細菌16S rRNA PCR 擴增及MiSeq 高通量測序
參考蔡宏宇等方法進行樣品細菌16S rRNA PCR擴增及MiSeq 高通量測序[19]。擴增體系總體積為20 μL,包括 5×PCR 緩沖液 4 μL,dNTP mix 2 μL,帶有 7個核苷酸標簽(barecode)的正向引物338F 和反向引物806R 各 0.8 μL,rTaq 酶 0.4 μL,模板 10 ng,無菌超純水補至 20 μL。擴增程序為:95℃預變性 3 min,95℃變性 30 s,55℃退火 30 s,72℃延伸 45 s,循環 30 次,72℃終延伸10 min。用1.0 %瓊脂糖凝膠電泳檢測PCR 擴增產物,寄至上海美吉生物醫藥科技進行Illumina MiSeq PE300 平臺進行高通量測序。
1.2.4 序列拼接及質量控制
高通量測序數據成功下機后,參考董蘊等方法對序列進行質控和拼接[20]:根據成對序列之間的重疊關系,將雙端序列拼接成一條序列并去除不合格序列,從而完成序列校正。利用QIIME 分析軟件,將相似度〉97%以上的序列歸類為一個操作分類單元(operational taxonomic unit,OTU),從而確定各序列對應微生物的分類學地位及相對含量,同時根據Chao 1 指數和Shannon指數對各樣品的微生物豐富度和多樣性進行分析。本研究將在4個樣品中均存在的OTU 定義為核心OTU。
1.3 數據處理
使用Origin 8.5 軟件對稀釋曲線及香農指數曲線作圖,同時對麥曲樣品中優勢細菌屬進行柱狀圖的繪制。各特征條帶序列系統發育樹由Bio Edit 軟件和MEGA 7.0 軟件共同繪制。基于OTU 水平的Venn 圖由在線網站(http://bioinfogp.cnb.csic.es/tools/venny/index.html)進行繪制。
2 結果與分析
2.1 麥曲細菌DGGE電泳圖譜及條帶測序分析
本試驗首先利用PCR-DGGE 技術對麥曲樣品中的細菌多樣性進行了電泳分析,麥曲中細菌PCRDGGE 圖譜結果如圖1所示。

圖1 麥曲中細菌PCR-DGGE 圖譜Fig.1 PCR-DGGE analysis of bacteria in wheat Qu
由圖1可知,共有8個條帶在電泳圖中明顯表現出來。條帶 2、3、4、5、6 和 7 均存在于麥曲樣品中,而條帶1 僅存在于MQ4中,條帶8 存在于MQ1 和MQ4中,由此可知各樣品細菌組成存在差異。將各條帶切膠回收進行PCR 擴增、測序、比對及系統發育樹繪制,其結果如表2所示。
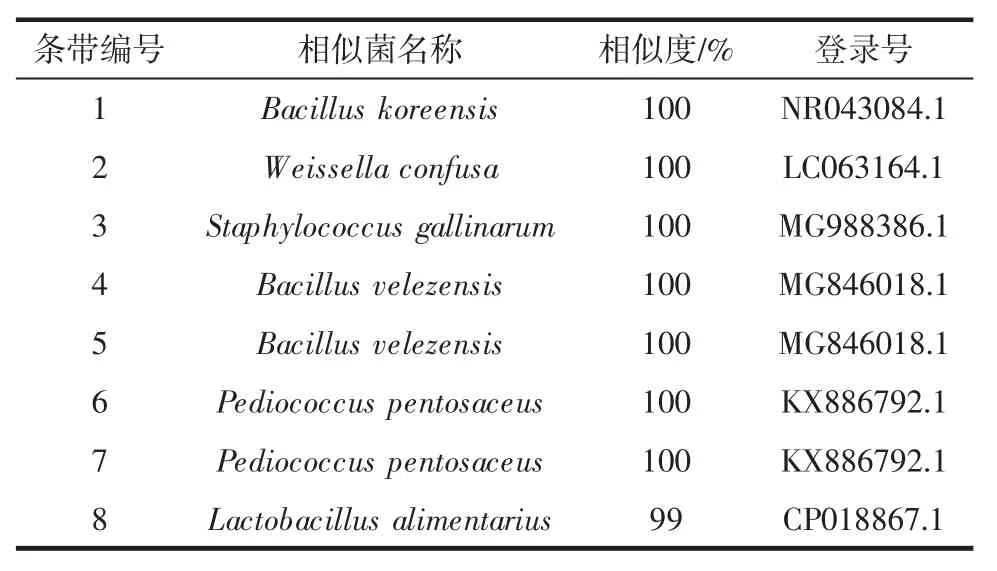
表2 麥曲DGGE 指紋圖譜中條帶比對結果Table 2 The blast results of bands in DGGE fingerprint of wheat Qu
由表2可知,條帶 1、2 和 3 分別與 Bacillus koreensis(韓國芽孢桿菌)、Weissella confusa(融合魏斯氏菌)和Staphylococcus gallinarum(雞葡萄球菌)相似度達 100%,條帶 4 和 5 與菌 Bacillus velezensis(貝萊斯芽孢桿)相似度達100%,同時條帶6 和7 與Pediococcus pentosaceus(戊糖片球菌)相似度達100%,而條帶8 與Lactobacillus alimentarius(食品乳桿菌)相似度為99%。系統發育樹見圖2。
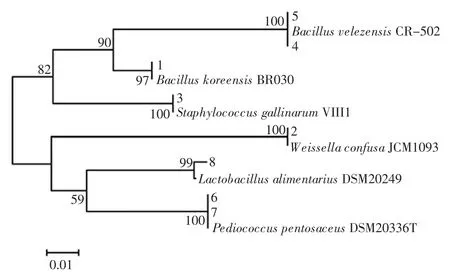
圖2 系統發育樹Fig.2 Phylogenetic tree
由圖2可知,系統發育樹被分為兩大分支,說明各分支上聚集的菌株與數據庫中的模式菌有較強的親緣關系。其中條帶 1、3、4 和 5 聚為一類,而條帶 2、6、7和8 聚為一類。在DGGE 指紋圖譜中不同條帶代表不同的細菌種類,體現了樣品微生物的豐富程度,條帶越亮或越粗表示其代表的種群生物量越大[21]。由此可見,麥曲中的細菌主要隸屬于Bacillus(芽孢桿菌)、Weissella(魏斯氏菌)和Pediococcus(片球菌)。薛景波利用16S rDNA 分子學方法對黃酒接種生麥曲發酵過程中微生物群落的變化研究發現Weissella confusa(融合魏斯氏菌)和Enterobacter cloacae(阿氏腸桿菌)的豐度較高,并被確定為優勢細菌[22]。
2.2 序列豐富度及多樣性分析
本試驗進一步將各麥曲樣品進行MiSeq 高通量測序,4個樣品共產生220 707 條序列。根據兩步UCLUST法對所有序列進行分析,首先經100%相似度進行聚類,共得到73 688 條代表性序列,依據97%相似度聚類后得到5 726個OTU,平均每個樣品1 431個OTU。樣品16S rRNA 測序情況及各分類地位數量如表3所示。

表3 樣品16S rRNA 測序情況及各分類地位數量Table 3 16S rRNA read counts and the number of identifiable units on different taxonomical levels
由表3 亦可知,MQ1 樣品中細菌群落豐富度最大,而MQ2 樣品中細菌群落多樣性最高。稀釋曲線可以直接反映測序數據量的合理性,香農曲線是用來衡量個樣品之間微生物群落的差異性[23]。本研究進一步通過稀疏曲線和香農指數曲線對測序深度是否滿足生物信息學分析進行了評價,其結果如圖3所示。
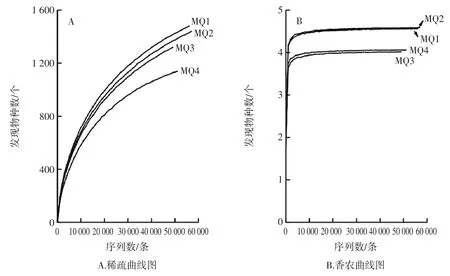
圖3 稀疏曲線圖和香農曲線圖Fig.3 Rarefaction curve and Shannon index curve
由圖3可知,隨著測序序列數的增加,雖然會有新的細菌種系型可能會被發現,但是微生物多樣性卻處于飽和狀態,由此可知上述測序量可以滿足生物信息學分析。
2.3 基于不同分類學地位細菌群落相對含量分析
納入本研究的4個麥曲樣品,共產生220 707 條序列,通過同源性比對將其鑒定為12個門、29個綱、49個目、96個科和170個屬,只有0.94%和5.64%的序列不能鑒定到門和屬水平。在門水平上,Firmicutes(硬壁菌門)、Proteobacteria(變形菌門)和Actinobacteria(放線菌門)為優勢細菌門,其中Firmicutes(硬壁菌門)平均相對含量最高,而Actinobacteria(放線菌門)平均相對含量最低,分別為87.97%和0.94%,由此可知隸屬于Firmicutes(硬壁菌門)的細菌為麥曲樣品中的優勢細菌。同時各樣品中硬Firmicutes(壁菌門)相對含量分別為 88.13%、89.13%、90.04 和 84.48%,Proteobacteria(變形菌門)相對含量分別為9.17%、8.93%、7.18%和10.72 %,而Actinobacteria(放線菌門)相對含量為1.13%、0.57%、1.34%和1.58%,由此可知雖然上述各細菌門均存在于各樣品中,但是相對含量卻存在差異性。本研究進一步將相對含量〉1%的細菌屬進行分析,結果如圖4所示。
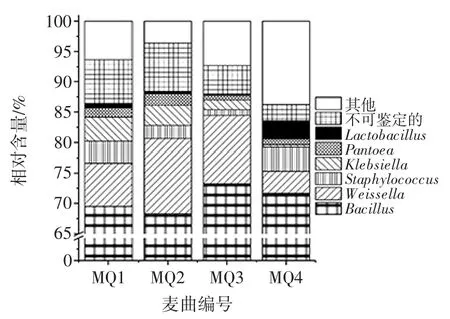
圖4 麥曲樣品中優勢細菌門相對含量分析Fig.4 Analysis of relative abundance of dominant bacterial in wheat Qu samples at the phylum level
由圖4可知,優勢細菌屬分別為Bacillus(芽孢桿菌屬)、Weissella(魏斯氏屬)、Staphylococcus(葡萄球菌)、Klebsiella(克雷伯菌屬)、Pantoea(泛菌屬)和Lactobacillus(乳桿菌屬),其平均相對含量分別為70.70%、8.55 %、2.67 %、2.29 %、1.32 %和 1.07 %。利用 Illumina MiSeq 測序平臺,劉蕓雅對紹興黃酒麥曲及黃酒發酵過程中細菌多樣性進行了分析,結果發現麥曲中優勢菌為Bacillus(芽孢桿菌屬)和Saccharomyces(糖多孢菌屬),同時Bacillus(芽孢桿菌屬)、Staphylococcus(葡萄球菌屬)和Lactobacillus(乳桿菌屬)為黃酒發酵過程中的優勢細菌屬[24]。利用傳統微生物學方法,張中華對熟麥曲中的細菌進行了培養、分離和鑒定,結果發現 Bacillus(芽孢桿菌屬)、Staphylococcus(葡萄球菌屬)和Pantoea(泛菌屬)細菌存在于熟麥曲中[25]。任清通過純培養和分子生物學方法對北宗黃酒麥曲中的細菌進行鑒定,結果發現Bacillus(芽孢桿菌屬)為麥曲中的優勢細菌[26],該報道與本研究結論一致。
本研究進一步將每個樣品中的OTU 數量及序列進行統計,結果如圖5所示。
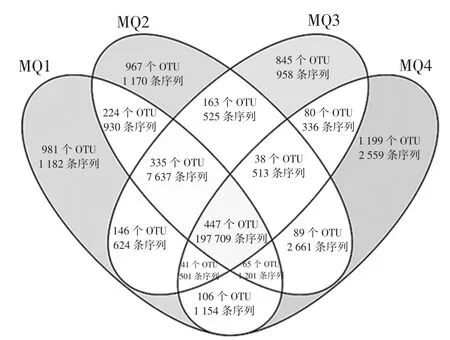
圖5 基于OTU 水平的Venn 圖Fig.5 Venn diagram based on OTU level
由圖5可知,4個樣品中共發現5 726個OTU,而僅存在1個樣品中的OTU 為3 992個,占OTU 總數的69.72%,序列數為5 869 條。同時出現2 次和3 次的OTU 分別為808個和479個,分別占 OTU 總數的14.11%和8.37%,包含序列數分別為6 230 條和9 852條。值得一提的是在4個樣品中共發現447個核心OTU,占OTU 總數的7.80%,包含197 709 條序列。進一步分析發現,在核心OTU中有7個OTU 的平均相對含量〉1%,結果如圖6所示。
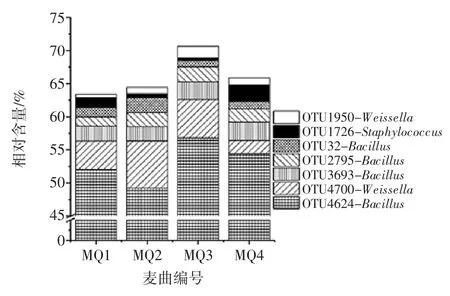
圖6 平均相對含量>1%核心OTU 比較分析Fig.6 Comparative analysis of average relative abundance more than 1%of core OTU
由圖6可知,平均相對含量〉1%的核心OTU 分別為OTU4624、OTU4700、OTU3693、OTU2795、OTU32、OTU1726 和OTU1950,其平均相對含量分別為54.44%、1.97%、2.78%、1.99%、1.10%、2.56%和1.03%。進一步將上述7個核心OTU 序列進行Blast 比對發現OTU4624、OTU2795 和 OTU3693 隸屬于 Bacillus(芽孢桿菌屬),OTU4700 和 OTU1950 隸屬于 Weissella(魏斯氏屬),而OTU 隸屬于Staphylococcus(葡萄球菌屬)。
3 結論
本研究以麥曲為研究對象,采用PCR-DGGE 與MiSeq 高通量測序技術相結合的手段對其細菌多樣性進行評價分析。結果發現,麥曲中的細菌組成具有較高的多樣性,主要為硬壁菌門、變形菌門和放線菌門,其中隸屬于硬壁菌門的芽孢桿菌屬與魏斯氏屬是為優勢菌屬。通過本試驗的開展,為后續麥曲中微生物資源的挖掘提供了理論支持。
