高效利用棉籽糖菌種的篩選及酶學(xué)性質(zhì)研究
2019-06-06 06:51:56陳磊劉羽彤童群義
食品與發(fā)酵工業(yè) 2019年10期
陳磊,劉羽彤,童群義
(江南大學(xué) 食品學(xué)院,江蘇 無錫,214122)
我國作為一個農(nóng)業(yè)大國,擁有大量的棉粕和豆粕資源,且棉籽與豆粕具有售價低、氨基酸組成合理等特點,是我國飼料原料的首選[1-2]。但由于棉籽和大豆中含有的抗?fàn)I養(yǎng)因子——低聚糖,尤其是棉籽糖的含量較高,嚴(yán)重的影響了應(yīng)用及營養(yǎng)因子的吸收利用。棉籽糖(raffinose)是一種由一分子的半乳糖、葡萄糖、果糖組成的功能性低聚糖,非還原糖,又稱為蜜三糖、棉實糖、棉子糖,分子式為C18H32O16,相對分子質(zhì)量為504.46。棉籽糖在熱和酸的環(huán)境中有較強(qiáng)的穩(wěn)定性[3],很難通過常規(guī)理化方法去除。
α-半乳糖苷酶(α-galactosidase)又稱密二糖酶,能水解通過α-1, 6-鍵結(jié)合的末端半乳糖殘基類物質(zhì)(蜜二糖、棉籽糖、糖鞘脂、糖蛋白等)[4]。由于人和單胃動物體內(nèi)不含有α-半乳糖苷酶,所以無法消化棉籽糖,棉籽糖在進(jìn)入大腸后被發(fā)酵產(chǎn)酸產(chǎn)氣,從而引起脹氣、嘔吐、腹瀉等不良現(xiàn)象。利用α-半乳糖苷酶可將結(jié)構(gòu)復(fù)雜的低聚半乳糖分解為小分子糖而成被利用,從而有效避免脹氣等[5-6]。
此外α-半乳糖苷酶水解半乳甘露聚糖的側(cè)鏈促進(jìn)β-甘露聚糖酶水解[7],或通過轉(zhuǎn)糖苷合成α-低聚半乳糖及其他重要的新型糖苷化合物[8]。微生物中α-半乳糖苷酶常見于細(xì)菌和真菌中,僅有少數(shù)來自于古菌。近年用于基因克隆和酶活高效表達(dá)的菌種有BacteroidesfragilisNCTC9343[9]、Dictyoglomusthernaophilum[10]、Mesorhizobiumsp. JB07[11]、Porrtibactersp.HJ8[12]、NeosartoryafischeriP1[13]、TalaromycesleycettanusJCM12802[14]。本實驗旨在篩選出可以高效利用棉籽糖的菌種,并對其酶學(xué)性質(zhì)進(jìn)行研究,為含有棉籽糖原料中棉籽糖的消除應(yīng)用研究提供參考。
1 材料與方法
1.1 菌株
由飼料工廠附近土壤中篩選和實驗室保存的菌種。
1.2 材料與試劑
對硝基苯α-D-半乳糖苷、對硝基酚、棉籽糖:麥克林公司;其余試劑均為國藥集團(tuán)化學(xué)試劑有限公司提供。
1.3 儀器與設(shè)備
SJ-CJ-1FD超凈工作臺,蘇凈公司;GI80T滅菌鍋,致微(廈門)儀器有限公司;UV-2800a型分光光度計,尤尼柯(上海)儀器有限公司;One Shot 1532型超高壓破碎機(jī),Constant Systems LTD;FE28型pH計,梅特勒-托利多公司;ZQZY-70BF型雙層振蕩培養(yǎng)箱,上海知楚儀器有限公司;GH-400BC型恒溫培養(yǎng)箱,北京市永光明醫(yī)療儀器廠;RJ-TGL-1850R高速冷凍離心機(jī),無錫瑞江分析儀器有限公司。
1.4 培養(yǎng)基
LB培養(yǎng)基(g/L):蛋白胨10,牛肉膏3,NaCl 5。
YPD培養(yǎng)基(g/L):胰蛋白胨10,葡萄糖20,酵母膏10。
MRS培養(yǎng)基(g/L):蛋白胨10,牛肉膏10,酵母粉5,葡萄糖20,三水醋酸鈉5,吐溫80 1,檸檬酸三銨2.0,K2HPO42.0,MgSO4·7H2O 0.2,MnSO4·4H2O 0.05。
發(fā)酵培養(yǎng)基:將上述生長培養(yǎng)基中的葡萄糖用等量的棉籽糖取代。
1.5 方法
1.5.1 生物量的測定
取發(fā)酵液選擇2~3個連續(xù)的適宜稀釋濃度,每個稀釋度分別吸取200 μL均勻涂布于培養(yǎng)基平板上,37 ℃恒溫培養(yǎng)48 h后計數(shù),每個濃度做 3次平行。
1.5.2 棉籽糖降解能力研究
棉籽糖含量采用GB 5009.258—2016測定。
1.5.3 α-半乳糖苷酶活力測定方法
α-半乳糖苷酶酶活測定參考GARROAO等[15]的方法并做適當(dāng)修改。取適量發(fā)酵液,10 000×g離心15 min, 收集細(xì)胞沉淀。用檸檬酸鈉緩沖液洗滌細(xì)胞體,懸浮于1 mL緩沖液中,置于高壓細(xì)胞破碎機(jī)破壁,12 000×g離心20 min,上清液即為粗酶液。
將10 mmol/L pNPG溶液200 μL,檸檬酸緩沖液100 μL和粗酶液100 μL混合,37 ℃反應(yīng)15 min,加入3 mL Na2CO3緩沖液,于405 nm處測定吸光度[15]。
1.5.4 酶學(xué)性質(zhì)研究
1.5.4.1 α-半乳糖苷酶的最適pH和pH穩(wěn)定性
將粗酶液在不同的pH下,以pNPG法37 ℃測定不同pH值下的酶活。以測得的最高值為100%,其余數(shù)據(jù)折算。
將粗酶液在37 ℃,不同pH緩沖液中反應(yīng)2 h,以各pH值下反應(yīng)15 min時測得的酶活為基準(zhǔn)酶活(100%),其余的數(shù)據(jù)折算,繪制pH穩(wěn)定性曲線。
1.5.4.2 α-半乳糖苷酶的最適溫度和熱穩(wěn)定性
將粗酶液在不同的溫度下,以pNPG法最適pH值下測定不同溫度時的酶活。以測得的最高值為100%,其余數(shù)據(jù)折算。
將粗酶液在最適pH值時,不同溫度中反應(yīng)2 h,每30 min取樣以pNPG法測定酶活,以各溫度值時反應(yīng)15 min時測得的酶活為基準(zhǔn)酶活(100%),其余的數(shù)據(jù)折算,繪制熱穩(wěn)定性曲線。
1.5.4.3 不同金屬離子對酶活的影響
用濃度為 10 mmol/L的Zn2+、Cu2+、Mn2+、Ca2+、Fe2+、Fe3+、K+、Na+、Mg2+、Pb2+、Co2+、Ni2+、Ag+、Hg2+溶液分別與粗酶液以1:1的比例進(jìn)行混合,37 ℃保存1 h后,在最適條件下測定酶活。以未處理的酶液的酶活為100%計[16]。
1.5.4.4 粗酶液對蛋白酶的抗性研究
將粗酶液與1 g/L的胰蛋白酶、蛋白酶K、枯草桿菌蛋白酶A、堿性蛋白酶溶液按 10:1比例反應(yīng)30 min, 按pNPG法測定酶的剩余酶活。以未處理的酶液的酶活為100%計[17]。
2 結(jié)果與分析
2.1 生物量的測定
各菌種分別用無碳源培養(yǎng)基和棉籽糖為唯一碳源培養(yǎng)基分別培養(yǎng)48 h,梯度稀釋后選取3個適宜的稀釋度涂布平板,培養(yǎng)48 h后進(jìn)行計數(shù),結(jié)果如表1所示。
由表可知所有菌種都可利用棉籽糖進(jìn)行生長,凝結(jié)芽孢桿菌、屎腸球菌、糞腸球菌在棉籽糖培養(yǎng)基中比無碳源培養(yǎng)基中分別高出了45.33、38.24、31.58倍,說明3種菌可更好地利用棉籽糖進(jìn)行生長。
2.2 棉籽糖降解能力
以GB 5009.258—2016所述方法測定發(fā)酵結(jié)束后棉籽糖的剩余含量,結(jié)果如表2所示。

表2 不同菌種對棉籽糖的降解能力 單位:%(質(zhì)量分?jǐn)?shù))
由表可以看出,發(fā)酵48 h小時后各菌種均表現(xiàn)出對棉籽糖良好的降解能力,乳酸菌表現(xiàn)出優(yōu)于芽孢桿菌和酵母的分解能力。乳酸菌可分解85%以上的棉籽糖,屎腸球菌和糞腸球菌可以完全或基本完全分解棉籽糖。枯草芽孢桿菌和地衣芽孢桿菌分解能力低于乳酸菌,但凝結(jié)芽孢桿菌也表現(xiàn)出優(yōu)異的分解棉籽糖的能力,發(fā)酵48 h可分解98.95%的棉籽糖。
2.3 酶學(xué)性質(zhì)研究
由上述實驗可看出屎腸球菌、糞腸球菌、凝結(jié)芽孢桿菌可利用棉籽糖進(jìn)行良好的生長,并且3種菌都表現(xiàn)出優(yōu)異的分解棉籽糖的能力,基于以上結(jié)果,本部分實驗將進(jìn)一步對3種菌的粗酶液的酶學(xué)性質(zhì)進(jìn)行研究。
2.3.1 α-半乳糖苷酶的最適溫度和熱穩(wěn)定性
以pNPG法測定3種菌不同溫度時的酶活,結(jié)果如圖1所示。由圖1可知,凝結(jié)芽孢桿菌和屎腸球菌的最適溫度均為45 ℃,糞腸球菌的最適溫度為50 ℃。 隨著溫度升高酶活逐漸增加,當(dāng)達(dá)到最適溫度后酶活隨著溫度的升高而快速下降。

圖1 不同菌種的α-半乳糖苷酶的最適溫度Fig.1 Optimum temperature of α-galactosidase from different strains
在實際生產(chǎn)中對酶的熱穩(wěn)定性有著不同的要求,例如在飼料加工工藝中關(guān)鍵控制點調(diào)質(zhì)溫度一般為55~65 ℃,制粒和膨化溫度可達(dá)到70 ℃以上[16],且完整加工過程中物料會在高溫環(huán)境下保持較長的時間,所以酶的熱穩(wěn)定性對于酶的應(yīng)用至關(guān)重要。將3種粗酶液分別置于37、50、55、60、65 ℃下保持120 min,每30 min以pNPG法測定剩余酶活,結(jié)果如圖2所示。由圖可知:隨著加熱時間的延長酶活逐漸降低。糞腸球菌在37 ℃時酶活可保持較高水平,超過50 ℃后酶活嚴(yán)重下降,65 ℃, 120 min后基本失活。屎腸球菌在37和50 ℃酶活可保持在80%以上,55與60 ℃ 在120 min后酶活維持在45%以上,65 ℃時酶活僅剩20%。凝結(jié)芽孢桿菌在37和50 ℃時酶活可保持在90%以上,55 ℃時也可保持在近65%,在65 ℃ 時30 min仍可保持55%左右,但時間繼續(xù)增加后酶活嚴(yán)重下降,酶活在120 min時為18%。3種α-半乳糖苷酶都表現(xiàn)出優(yōu)于普通菌種[18-19]的熱穩(wěn)定性,但差于嗜熱菌NeosartoryafischeriP1[20]、Geobacillusstearotherrnophilus[21]、新阿波羅棲熱袍菌[22]所產(chǎn)α-半乳糖苷酶的熱穩(wěn)定性。

a-糞腸球菌;b-屎腸球菌;c-凝結(jié)芽孢桿菌圖2 α-半乳糖苷酶的熱穩(wěn)定性Fig.2 Thermal stability of α-galactosidase
2.3.2 α-半乳糖苷酶的最適pH和pH穩(wěn)定性
在37 ℃條件下,以pNPG法測定3種α-半乳糖苷酶在不同pH緩沖液時的酶活,結(jié)果如圖3所示。
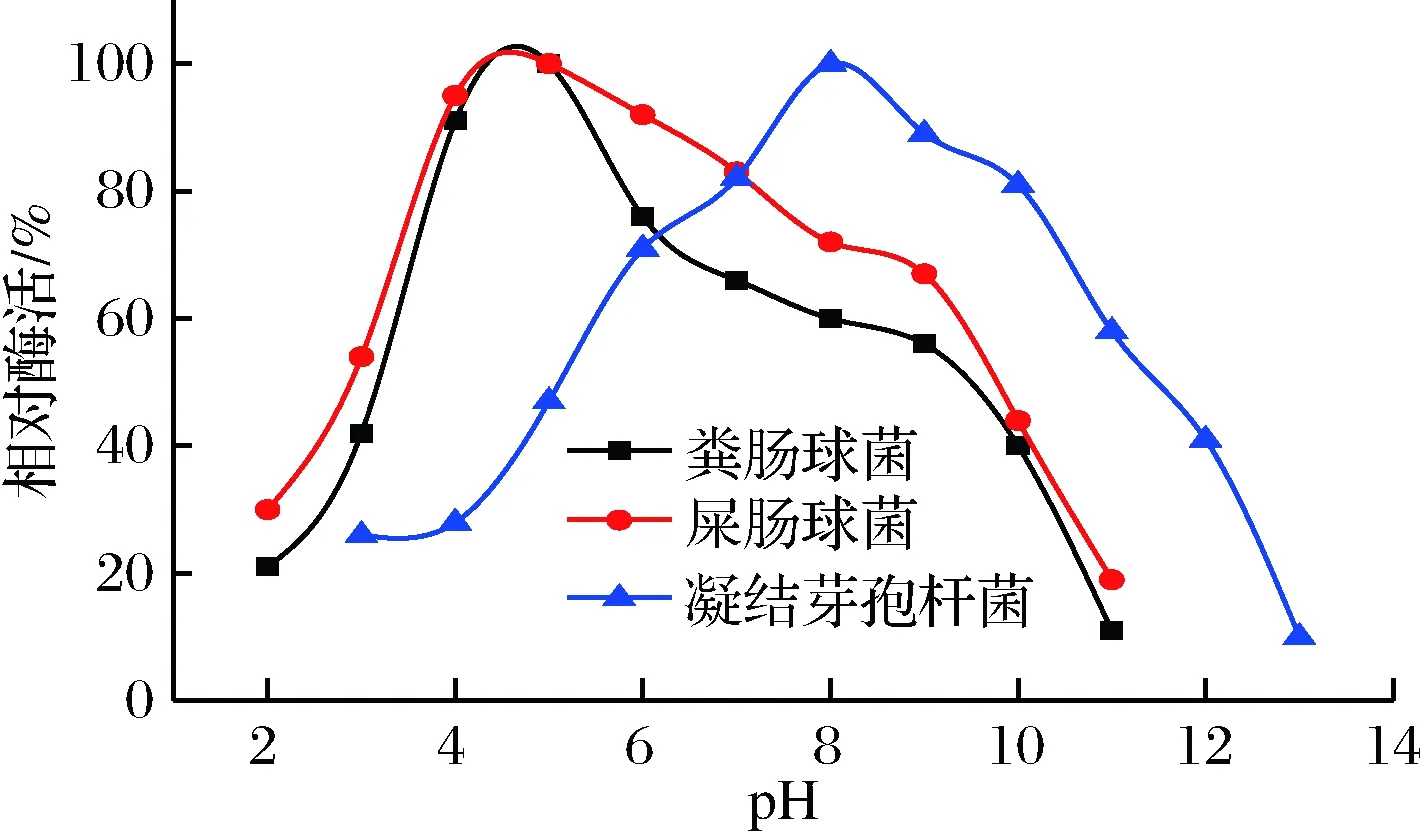
圖3 不同菌種的α-半乳糖苷酶的最適pHFig.3 Optimum pH of α-galactosidase from different strains
由圖3可以看出,糞腸球菌和屎腸球菌α-半乳糖苷酶在pH值為4~8具有較高的酶活,均可保持60%以上,最適pH值為5。凝結(jié)芽孢桿菌α-半乳糖苷酶最適pH值為8,其在6~11具有>60%的較高的酶活。
將粗酶液在37 ℃各pH值條件下反應(yīng)120 min,以pNPG法測定3種酶在不同pH緩沖液時的剩余酶活,以各pH值時反應(yīng)15 min時測得的酶活為基準(zhǔn)酶活(100%),結(jié)果如圖4所示。
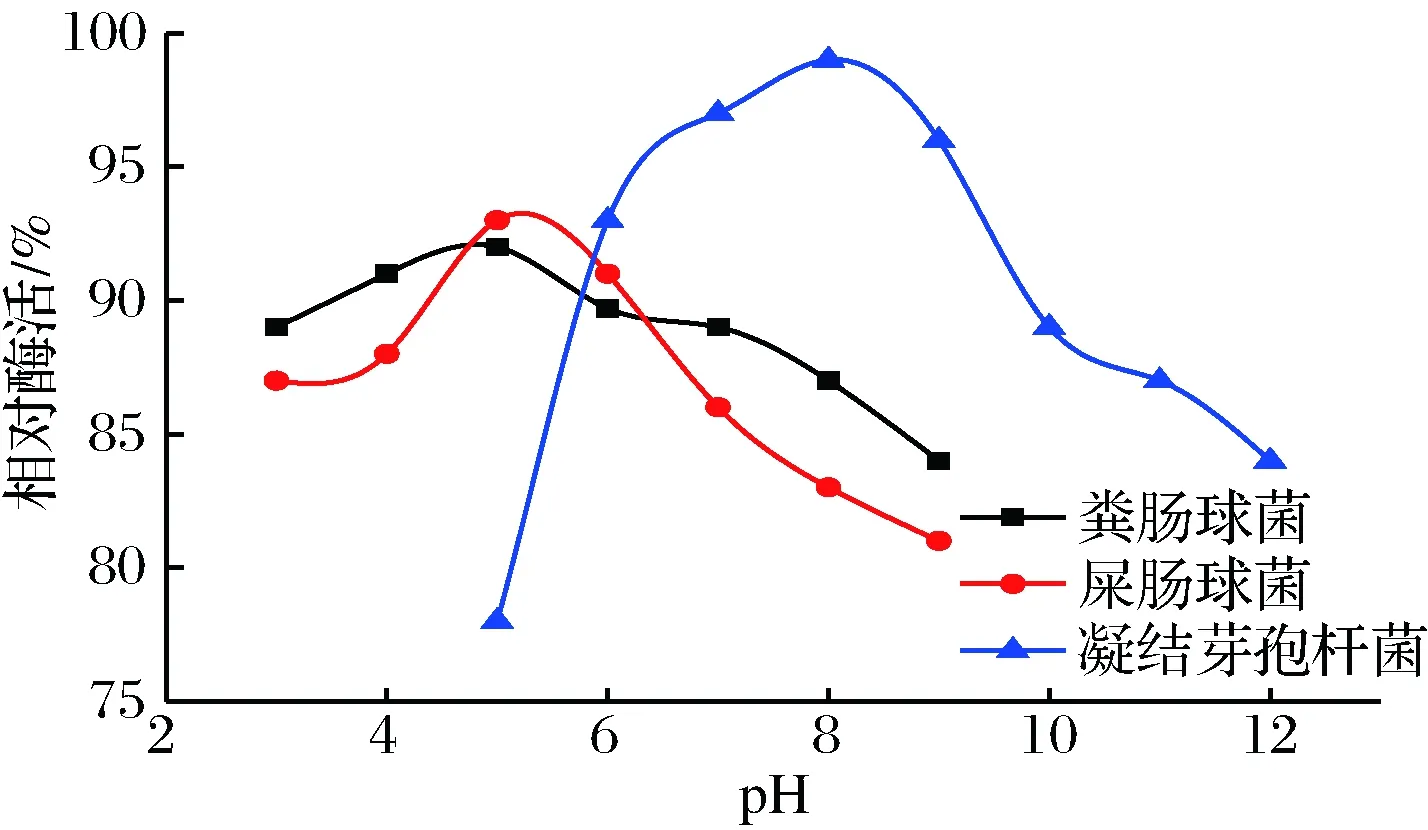
圖4 不同菌種的α-半乳糖苷酶的pH穩(wěn)定性Fig.4 pH stability of α-galactosidase from different strains
由圖4可知, 3種α-半乳糖苷酶在最適pH處都具有最高的穩(wěn)定性。這與董巖巖[16]、DU[23]、HU[24]已報道的α-半乳糖苷酶的pH穩(wěn)定性的結(jié)論相同,但與葉雪飛等[25]的結(jié)論相反。屎腸球菌和糞腸球菌α-半乳糖苷酶在pH 3~9都可保持在80%以上,但在最適pH處屎腸球菌α-半乳糖苷酶酶活高于糞腸球菌α-半乳糖苷酶。凝結(jié)芽孢桿菌α-半乳糖苷酶在pH 6~9具有90%以上的酶活,尤其是在最適pH處具有98%的相對酶活。
2.3.3 金屬離子對α-半乳糖苷酶酶活的影響
由表3可以看出,不同金屬離子對α-半乳糖苷酶有著不同的影響,且同一種金屬離子對不同的α-半乳糖苷酶的影響程度也不盡同。

表3 金屬離子對α-半乳糖苷酶酶活的影響Table 3 Effect of metal ions on α-galactosidase activity
注:數(shù)據(jù)由3個獨立試驗的均值± SD所示。
大部分金屬離子會對3種不同的α-半乳糖苷酶產(chǎn)生輕微的抑制作用,Zn2+離子對3種α-半乳糖苷酶有輕微的促進(jìn)作用,Mn2+對α-半乳糖苷酶酶活具有明顯的促進(jìn)作用。Ag+、Hg2+可完全抑制3種不同的α-半乳糖苷酶的酶活,這也與之前的報導(dǎo)結(jié)果[24,26-27]相一致。
2.3.4 α-半乳糖苷酶對蛋白酶的抗性研究
將粗酶液與胰蛋白酶、蛋白酶K、枯草桿菌蛋白酶A、糜蛋白酶溶液按 10∶1比例反應(yīng)30 min,按pNPG法測定酶的剩余酶活。由圖5可看出,糞腸球菌α-半乳糖苷酶對蛋白酶K有較好的抗性,但對其余蛋白酶的抗性較差。屎腸球菌α-半乳糖苷酶對糜蛋白酶的抗性較差為72%,其余3種酶抗性較好均>80%。凝結(jié)芽孢桿菌α-半乳糖苷酶對4種蛋白酶均表現(xiàn)出優(yōu)異的抗性,剩余酶活均>89%。

圖5 蛋白酶對α-半乳糖苷酶酶活的影響Fig.5 Effect of protease on α-galactosidase activity
3 結(jié)論
從飼料工廠土壤中和實驗室已保存菌種中篩選出了3種可高效利用棉籽糖的菌種,分別是屎腸球菌、糞腸球菌和凝結(jié)芽孢桿菌。
對粗酶液的酶學(xué)性質(zhì)研究表明:屎腸球菌α-半乳糖苷酶最適溫度為45 ℃,在55 ℃內(nèi)有較高的熱穩(wěn)定性,對4種蛋白酶有較好的抗性。糞腸球菌α-半乳糖苷酶最適溫度為50 ℃,對蛋白酶K有較好的抗性。屎腸球菌α-半乳糖苷酶與糞腸球菌α-半乳糖苷酶最適pH值均為5,在pH 3~9可保持較高酶活。凝結(jié)芽孢桿菌α-半乳糖苷酶最適溫度為45 ℃,最適pH為8,在55 ℃內(nèi)和pH 6~12可保持較高酶活,對4種蛋白酶有優(yōu)異的抗性。Mn2+對3種α-半乳糖苷酶酶活具有明顯的促進(jìn)作用,Ag+、Hg2+可完全抑制其酶活。
本實驗篩選到的3種菌種可為飼料、豆奶等中棉籽糖的消除提供理論基礎(chǔ)。下階段將對粗酶液進(jìn)行高效表達(dá)并分離純化得到高活性的α-半乳糖苷酶,并探究其實際應(yīng)用效果。
