富馬酸鈉對熒光假單胞菌群體感應現象及其腐敗活性的抑制作用
2019-07-01 07:50:16孫曉佳李婷婷赫彬彬梅永超王當豐勵建榮
食品科學 2019年11期
孫曉佳,李婷婷*,赫彬彬,梅永超,王當豐,謝 晶,勵建榮,*
(1.渤海大學食品科學與工程學院,生鮮農產品貯藏加工及安全控制技術國家地方聯合工程研究中心,遼寧省食品安全重點實驗室,遼寧省高等學校生鮮食品產業技術研究院,遼寧 錦州 121013;2.大連民族大學生命科學學院,遼寧 大連 116600;3.上海海洋大學食品學院,上海 201306)
群體感應(quorum sensing,QS)是一種細菌間信息交流的通訊系統,細菌通過自身合成并釋放信號分子感知環境變化,當信號分子濃度達到一定閾值時開啟細胞密度依賴的特定基因表達,進而調控細菌的生物行為,如生物發光、生物被膜形成、毒力因子表達及浮游等[1-2]。微生物的生長代謝是導致水產品腐敗的主要原因之一,其QS機制在水產品的腐敗過程中起著至關重要的作用。本實驗室從腐敗大菱鲆中分離得到一株熒光假單胞菌(Pseudomonas fluorescens,PF),作為水產品低溫貯藏過程中的優勢腐敗菌(specific spoilage organism,SSO),其腐敗特性受N-酰基高絲氨酸內酯(N-acyl-homoserine lactone,AHLs)介導的QS調控[3-5]。
群體感應抑制劑(quorum sensing inhibitors,QSIs)指對細菌間信息交流具有阻礙作用,能干擾或抑制QS現象,且對細菌正常生命活動不造成破壞的物質[6]。其作用機制主要有以下3 種途徑[7-9]:干擾信號分子合成,如抑制信號分子合成酶活性或消除底物等合成過程;降解信號分子,使之無法達到發揮調控作用的閾值濃度;產生信號分子類似物與受體蛋白競爭性結合。因此,利用QSI干擾細菌QS系統、降低腐敗菌致腐能力成為新的研究方向。研究發現阿魏精油作為一種新型的QSI可顯著抑制紫色桿菌的QS現象[10]。Chow等[11]報道了水楊酸對熒光假單胞菌QS現象具有抑制作用,可有效降低其毒力效應。目前已從藥用植物、果蔬、海洋藻類中分離出具有抑制QS的天然活性物質,但其提取率低且操作繁瑣,未得到廣泛應用。而從現有食品添加劑中篩選QSI,具有成本低廉、便于合成、安全性較高等優點。
富馬酸鈉又稱富馬酸一鈉,易溶于水,常作為食品添加劑用于配制酒、飲料、罐頭和果凍等產品,是一種較安全的酸味劑。一些研究表明其具有良好的抗氧化、抑菌作用。在魚、貝類罐頭中添加富馬酸鈉可有效防止其黑變[12];肖爾卿等[13]發現經過富馬酸鈉與超高壓協同處理使草莓汁中H2S含量顯著減少;此外,富馬酸鈉對腌制蔬菜中的腐敗菌具有較強的抑制作用[14]。但目前,關于富馬酸鈉的研究主要集中在其作為食品添加劑改善食品風味方面,而作為QSI的研究卻鮮有報道。
因此,本實驗以富馬酸鈉為研究對象,探究富馬酸鈉對熒光假單胞菌QS現象的抑制效果,以期拓寬富馬酸鈉的應用范圍,為新型水產品防腐保鮮劑的研發提供新思路,豐富水產品保鮮理論。
1 材料與方法
1.1 菌株與試劑
熒光假單胞菌分離自腐敗大菱鲆,由渤海大學食品科學研究院保藏。紫色桿菌CV026(Chromobacterium violaceum CV026)為C. violaceum ATCC 31532的mini-Tn5突變體,自身不產AHLs。當存在外源AHLs時,可產生特征性紫色菌素,對酰基側鏈長度為C4~C8的AHLs敏感程度不同,具有卡那霉素抗性,活化時需添加20 μg/mL卡那霉素,由渤海大學食品科學研究院保藏。
富馬酸鈉 北京索萊寶生物科技有限公司;LB肉湯青島高科園海博生物技術有限公司;甲醇 天津市津東天正精細化學試劑廠;乙酸乙酯、正丁醇、醋酸異戊酯、十二烷基硫酸鈉 天津永晟精細化工有限公司;結晶紫 天津致遠化學試劑有限公司;卡那霉素、6 種AHLs信號分子標準品(C4-HSL、C6-HSL、C8-HSL、C10-HSL、C12-HSL、C14-HSL) 美國Sigma公司。
1.2 儀器與設備
SW-CJ-2FD超凈工作臺 蘇凈集團蘇州安泰空氣技術有限公司;Biofuge Stratos臺式高速離心機 美國Thermo Fisher公司;MLS-3030CH立式壓力滅菌鍋廣州三洋電機有限公司;MS105UD電子分析天平瑞士梅特勒-托利多儀器有限公司;imark酶標儀美國Bio-Rad公司;80i顯微鏡 日本尼康公司;7890N/5975氣相色譜-質譜聯用(gas chromatography-mass spectrometer,GC-MS)儀 美國Agilent公司。
1.3 方法
1.3.1 富馬酸鈉的最小抑菌濃度測定及QS抑制活性的初篩
參考Diao Wenrui等[15]的方法,將報告菌株CV026和熒光假單胞菌過夜培養至對數期(106CFU/mL),按體積比1∶100接種于LB肉湯中,28 ℃、160 r/min振蕩培養至OD595nm為1左右,用牛津杯打孔,加入不同質量濃度梯度(1.0~10.0 mg/mL)的富馬酸鈉,以無菌去離子水為空白對照,28 ℃靜置培養24~48 h后,觀察菌株生長情況。
報告菌株CV026過夜活化后,按體積比1∶100接種于LB肉湯中,28 ℃、160 r/min振蕩培養至OD595nm為1左右,與100 mL含有20 μg/mL C6-HSL信號分子的LB營養瓊脂混勻,用牛津杯打孔,加入200 μL質量濃度分別為0.5、1.0、1.5 mg/mL和2.0 mg/mL的富馬酸鈉,以無菌去離子水為空白對照,28 ℃靜置培養24~48 h,觀察紫色菌素的產生情況。
1.3.2 富馬酸鈉對紫色菌素產生及紫色桿菌生長的影響
參照Kato等[16]方法并稍作修改,將過夜活化后的報告菌株CV026按體積比1∶100接種于LB肉湯中,加入終質量濃度為0.5、1.0、1.5 mg/mL和2.0 mg/mL的富馬酸鈉,并添加20 μg/mL的C6-HSL信號分子,28 ℃、160 r/min振蕩培養至OD595nm為1左右。取300 μL菌液于無菌離心管中,加入150 μL質量分數10%十二烷基硫酸鈉,振蕩10 s,加入600 μL正丁醇,振蕩5 s,10 000 r/min離心5 min,取上清液200 μL加入96 孔板中,于595 nm波長處測定OD值,以無菌去離子水為空白對照。同時,為確定富馬酸鈉對CV026菌株生長能力的影響,以不加信號分子的實驗組作為對照,其余步驟同上。
1.3.3 富馬酸鈉對熒光假單胞菌生物被膜的影響
1.3.3.1 富馬酸鈉對熒光假單胞菌生物被膜形成量的影響
根據Rode等[17]方法將熒光假單胞菌過夜培養至對數期(106CFU/mL),按體積比1∶100接種于LB肉湯中,分裝至無菌離心管,每管1 mL,加入富馬酸鈉使其終質量濃度分別為0.5、1.0、1.5 mg/mL和2.0 mg/mL,以無菌去離子水為陰性對照,以20 μg/mL C6-HSL為陽性對照,每組3 次重復,28 ℃靜置培養48 h。待菌液密度測定結束后,傾倒菌液,無菌水漂洗浮游菌3~5 次,無菌風干燥35 min,0.001 g/mL結晶紫溶液染色15 min,無菌水沖洗5~7 次,加入體積分數33%冰醋酸充分溶解,取200 μL于96 孔板中,在595 nm波長處測定OD值。按下式計算生物被膜形成率。
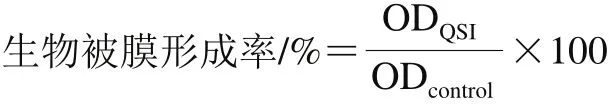
式中:ODcontrol為陰性對照組測得生物被膜的OD595nm;ODQSI為抑制劑處理組的OD595nm。
1.3.3.2 富馬酸鈉對熒光假單胞菌生物被膜形態的影響
載玻片(25.4 mm×76.2 mm,厚度1~2 mm)預處理:將載玻片清洗干凈,分別于無水乙醇中超聲30 min,去離子水中超聲30 min,烘干后滅菌備用。
將過夜活化后的熒光假單胞菌按體積比1∶100接種于LB肉湯中,取10 mL菌液加入無菌培養皿中,加入富馬酸鈉使其終質量濃度分別為0.5、1.0、1.5 mg/mL和2.0 mg/mL,將處理后的載玻片浸沒其中,28 ℃培養72 h。同時,以不添加富馬酸鈉為陰性對照組,添加C6-HSL信號分子為陽性對照組。取出載玻片后,無菌水漂洗3~5 次,適量甲醇固定15 min,0.02 g/mL結晶紫染色5 min,無菌水沖洗5~7 次,無菌風干燥固定,光學顯微鏡觀察并拍照記錄。
1.3.4 富馬酸鈉對熒光假單胞菌群集和泳動的影響
參考Zhang Juanmei等[18]方法制備群集與泳動培養基,將不同質量濃度的富馬酸鈉經0.22 μm濾膜過濾后,加入培養基混勻使其終質量濃度為0.5、1.0、1.5 mg/mL和2.0 mg/mL。取5 μL過夜活化的熒光假單胞菌菌懸液加入平板中央,待菌液吸收后28 ℃靜置培養48 h,以不添加富馬酸鈉為陰性對照,添加C6-HSL信號分子為陽性對照,每組3 次重復,期間觀察熒光假單胞菌的遷移情況。1.3.5 富馬酸鈉對熒光假單胞菌胞外蛋白酶活性的影響
根據Ponnuswamy等[19]的方法并稍作修改,按1.3.3節方法制備含有不同質量濃度(0.5、1.0、1.5 mg/mL和2.0 mg/mL)富馬酸鈉過夜活化后的熒光假單胞菌菌懸液,制作脫脂奶瓊脂平板,用牛津杯打孔,取200 μL菌液加入孔內,28 ℃靜置培養18~24 h,期間觀察其水解圈直徑。以不添加富馬酸鈉為陰性對照,以添加C6-HSL信號分子為陽性對照,每組3 次重復。熒光假單胞菌在生長過程中分泌胞外蛋白酶,使孔徑周圍出現明顯的水解圈,水解圈直徑的大小表示蛋白酶活性的高低。
1.3.6 GC-MS測定富馬酸鈉對熒光假單胞菌分泌AHLs的影響
將過夜活化后的熒光假單胞菌按體積比1∶100接種于LB肉湯中,加入富馬酸鈉,使其終質量濃度分別為0.5、1.0、1.5、2.0 mg/mL,以不添加富馬酸鈉為陰性對照,添加C6-HSL信號分子為陽性對照,28 ℃、160 r/min振蕩培養48 h。將培養后的菌液10 000 r/min離心10 min,取上清液,加入等量含體積分數0.1%冰乙酸的乙酸乙酯,得有機相,旋轉蒸發蒸干溶劑,條件設置為溫度35 ℃、真空度0.1 MPa,加入適量甲醇溶解,將所得AHLs粗提液于-20 ℃保存備用。
以甲醇為溶劑制備含有6 種AHLs標準品的混合標準品溶液,由GC-MS檢測確定各種標準品的保留時間。參考趙愛飛等[20]的方法設置GC-MS條件。
1.4 數據處理與分析
每組實驗均設有3 個平行,用SPSS 19.0軟件進行數據分析處理,Origin 8.5軟件進行繪圖處理。
2 結果與分析
2.1 富馬酸鈉的最小抑菌濃度測定及QS抑制活性初篩結果
采用平板法觀察富馬酸鈉質量濃度為1.0~10.0 mg/mL時CV026菌株的生長情況,確定了富馬酸鈉對CV026菌株的最小抑菌濃度為3.0 mg/mL。同時,相同質量濃度的富馬酸鈉對熒光假單胞菌不具有抑制作用(圖1A)。因此,選取亞抑菌濃度,即3.0 mg/mL以下質量濃度的富馬酸鈉進行后續實驗,實驗質量濃度設為0.5、1.0、1.5 mg/mL和2.0 mg/mL。圖1B為亞抑菌濃度下富馬酸鈉對CV026菌株QS活性的抑制效果,添加富馬酸鈉的孔徑周圍有乳白色、不透明的抑制圈出現,說明富馬酸鈉可以抑制CV026菌株特征性紫色菌素的產生。
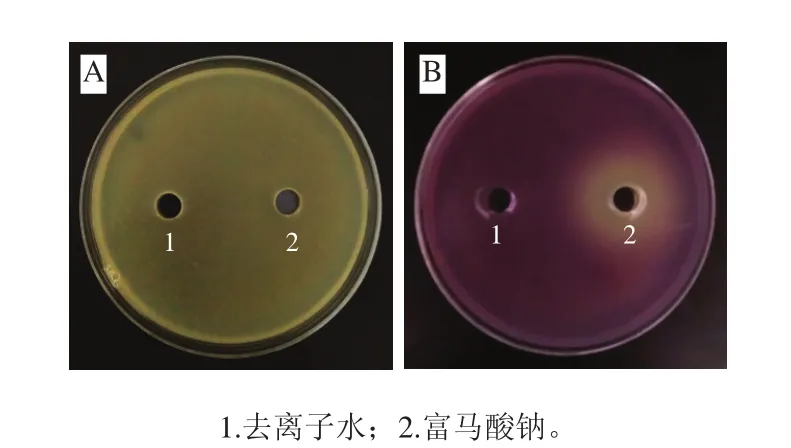
圖1 富馬酸鈉對熒光假單胞菌生長(A)及紫色桿菌CV026產紫色菌素能力(B)的抑制活性Fig. 1 Inhibitory effect of sodium fumarate on growth of Pseudomonas fluorescens (A) and violacein production of CV026 strains (B)
2.2 富馬酸鈉對紫色菌素產生及紫色桿菌生長的影響
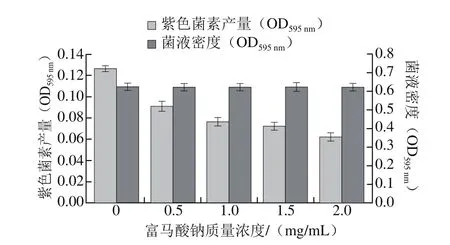
圖2 不同質量濃度富馬酸鈉對CV026菌株紫色菌素產量和菌液密度的影響Fig. 2 Effect of sodium fumarate on violacein production and bacterial density of CV026
由圖2可見,富馬酸鈉對CV026菌株QS現象的抑制效果。當富馬酸鈉質量濃度為2.0 mg/mL時,與對照組相比其紫色菌素產量明顯降低,其抑制率達50.95%;此外,隨著富馬酸鈉質量濃度逐漸增大,CV026菌株的菌液密度呈平穩狀態,說明其對CV026菌株的生長代謝能力沒有影響。由此可知,不同質量濃度的富馬酸鈉通過干擾細菌的QS系統,使得在C6-HSL信號分子存在的情況下,其紫色菌素的產生受到不同程度的抑制,且隨其質量濃度增加,這種抑制能力逐漸增強。
2.3 富馬酸鈉對熒光假單胞菌生物被膜的影響
2.3.1 富馬酸鈉對熒光假單胞菌生物被膜形成量的影響
生物被膜指附著在接觸物表面的細菌利用自身分泌多糖、纖維蛋白和脂蛋白等物質將菌體包裹形成的一種聚集狀膜樣物[21-22]。作為細菌保護自身的生長方式,其對環境變化的耐受力遠高于浮游菌,一旦在食品加工設備表面形成后極易對食品造成嚴重污染[23]。研究表明,由AHLs介導的QS系統對假單胞菌生物膜形成、附著及固定具有重要作用[24]。Zhang Ying等[25]報道了百里酚和水楊酸對熒光假單胞菌QS現象及生物被膜形成有較好的抑制效果。因此,本實驗探究不同質量濃度富馬酸鈉對熒光假單胞菌生物被膜的抑制作用。如圖3所示,當富馬酸鈉質量濃度為2.0 mg/mL時,其生物被膜形成率降低至31.82%;質量濃度為0.5 mg/mL時,其生物被膜形成率為84.05%,與陰性對照組相比均能抑制該菌生物膜的形成;而添加外源C6-HSL信號分子可促進其生物被膜形成。這表明,富馬酸鈉可能通過干擾AHLs介導的QS系統來抑制生物被膜的形成,且不影響其生長,與Zhang Ying等[25]的研究結果相符合。
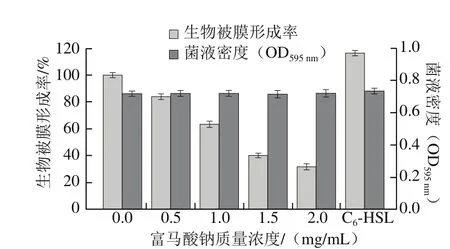
圖3 不同質量濃度富馬酸鈉對熒光假單胞菌生物被膜形成率的影響Fig. 3 Effect of sodium fumarate on biofilm formation rate of Pseudomonas fluorescens
2.3.2 富馬酸鈉對熒光假單胞菌生物被膜形態的影響
由圖4可見,陰性對照組有大量菌體相連形成密集的紫色膜狀結構,且陽性對照組有局部菌體聚集成具有層次感的深紫色團狀結構;而處理組隨富馬酸鈉質量濃度增加菌體數量明顯減少,部分菌體相連,呈稀疏的網狀結構。這說明富馬酸鈉對熒光假單胞菌生物被膜形成具有抑制作用,由此推測富馬酸鈉可能干擾了菌體聚集,從而減少細菌微菌落和生物被膜的形成。
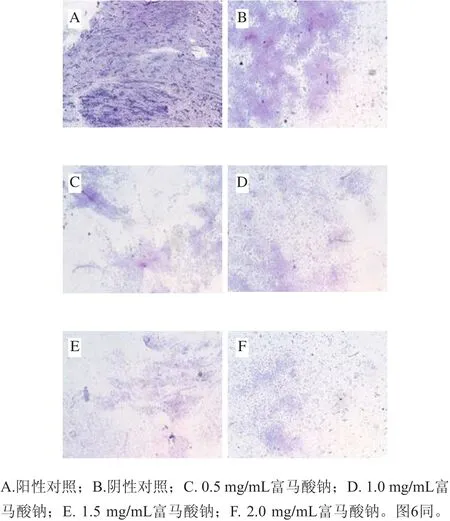
圖4 富馬酸鈉對熒光假單胞菌生物被膜影響的光學顯微鏡圖(×10)Fig. 4 Optical microscopic images showing the effect of sodium fumarate on biofilm formation of Pseudomonas fluorescens (× 10)
2.4 富馬酸鈉對熒光假單胞菌群集與泳動的影響
群集與泳動是由細菌鞭毛介導的兩種遷移方式[26],其不同之處在于前者是群體行為,指細菌在液體培養基內或固體培養基表面的遷移情況,后者則是一種個體行為,指在固體培養基表面或半固體培養基上的遷移情況[27]。細菌鞭毛介導的運動特性與初期黏附至宿主表面及沿著表面的遷移相關聯,并且其在后期形成生物被膜的過程中起關鍵作用[28]。圖5為不同質量濃度富馬酸鈉對熒光假單胞菌群集與泳動的抑制效果,與陰性對照組相比,陽性對照組運動性顯著增強,這表明外源AHLs可以促進該菌群集和泳動運動特性。隨著富馬酸鈉質量濃度的增加,其遷移能力呈濃度依賴性下降。這與上述富馬酸鈉對該菌生物被膜的抑制作用呈現出一定相關性,推測富馬酸鈉可能干擾了熒光假單胞菌黏附至接觸面的能力,從而減少生物被膜的形成。研究表明,肉桂醛能通過影響銅綠假單胞菌的群集和泳動運動性來減少生物被膜形成[29]。Lou Zaixiang等[30]也發現牛蒡葉成分可以減弱銅綠假單胞菌的群集運動能力,從而達到抑制生物被膜形成的目的,與本研究結果相似。
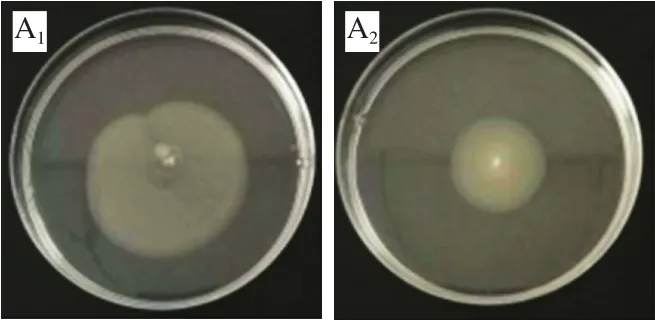
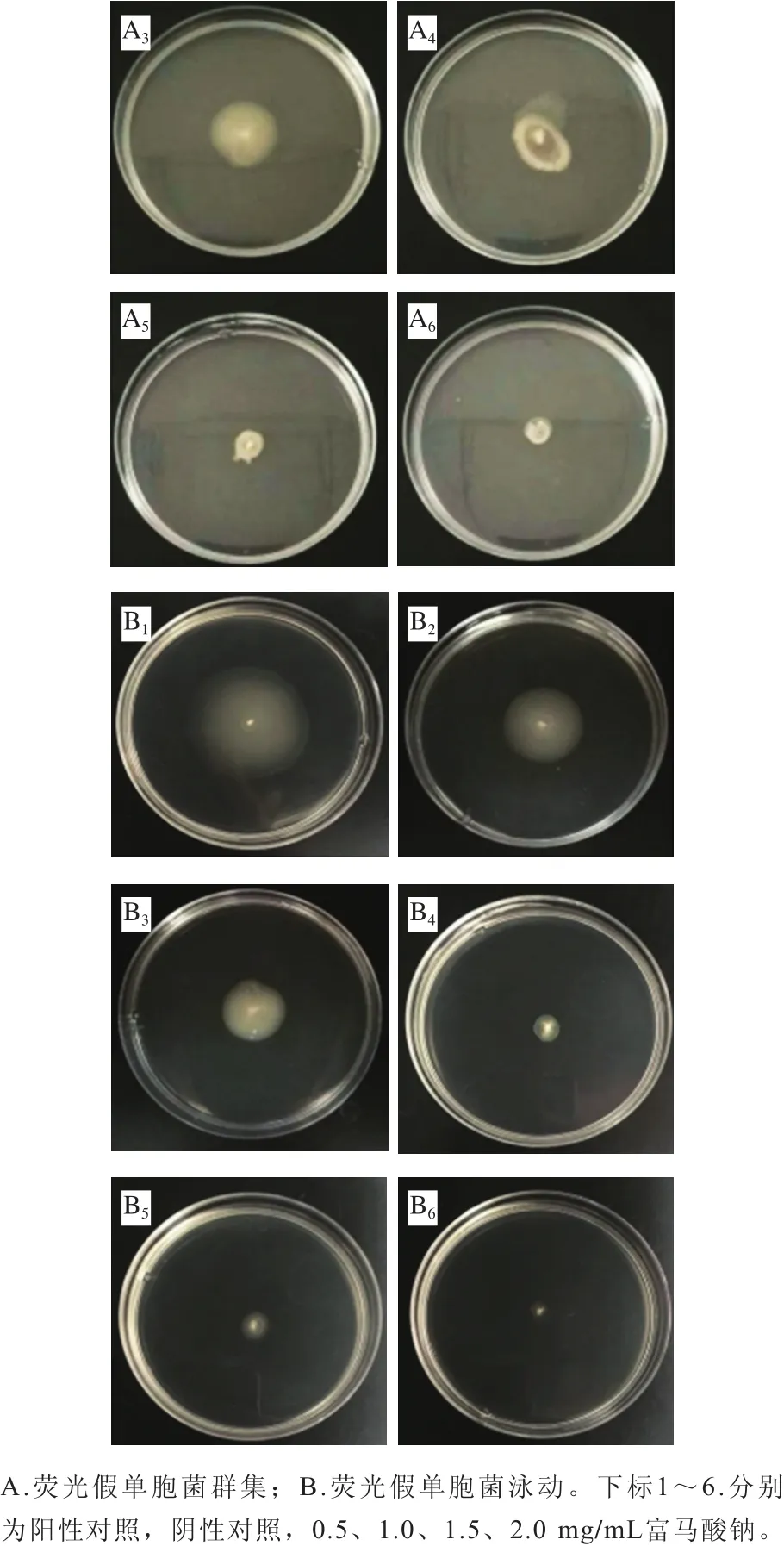
圖5 富馬酸鈉對熒光假單胞菌群集、泳動運動的影響Fig. 5 Effect of sodium fumarate on swarming and swimming motility of Pseudomonas fluorescens
2.5 富馬酸鈉對熒光假單胞菌胞外蛋白酶活性的影響
熒光假單胞菌可利用自身分泌的胞外蛋白酶將水產品中蛋白質及游離氨基酸等物質降解,產生多種腐敗代謝物以促進菌體生長代謝,進而使水產品風味和品質發生劣變[31]。相關研究表明,熒光假單胞菌所分泌的胞外蛋白酶受AHLs介導的QS系統調控[32]。Lokender等[33]發現姜油酮能通過干擾信號分子受體蛋白來抑制銅綠假單胞菌QS系統,從而降低蛋白酶活性。因此,本實驗探究不同質量濃度富馬酸鈉對熒光假單胞菌胞外蛋白酶活性的抑制作用。圖6、7顯示了富馬酸鈉對熒光假單胞菌胞外蛋白酶活性的影響。結果表明,富馬酸鈉對該菌蛋白酶活性具有明顯抑制作用,呈濃度依賴性增強;且外源C6-HSL信號分子的添加刺激該菌分泌AHLs,增強其蛋白酶活性,因此陽性對照組的水解圈直徑最大,達21.09 mm,與Lokender等[33]報道的結果一致。
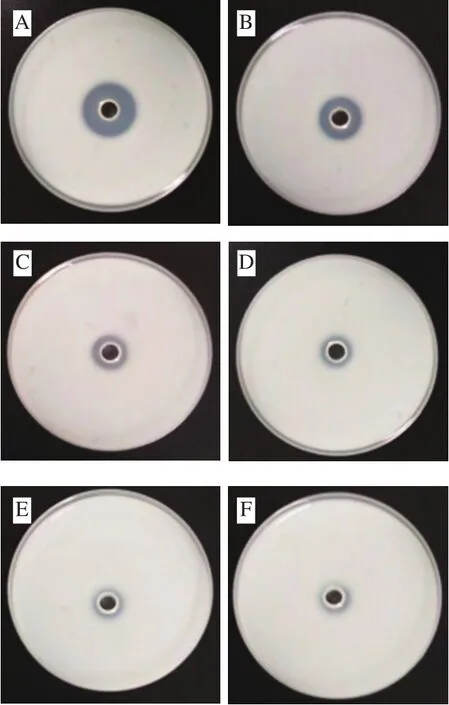
圖6 富馬酸鈉對熒光假單胞菌胞外蛋白酶活性的影響Fig. 6 Effect of sodium fumarate on extracellular protease activity of Pseudomonas fluorescens
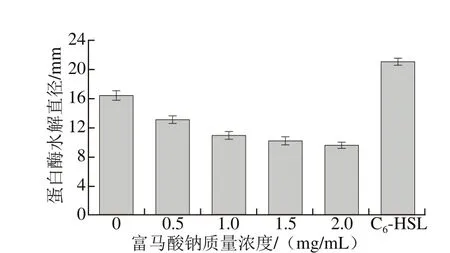
圖7 富馬酸鈉對熒光假單胞菌胞外蛋白酶活性的影響Fig. 7 Effect of sodium fumarate on extracellular protease activity of Pseudomonas fluorescens
2.6 GC-MS定量分析富馬酸鈉對熒光假單胞菌分泌AHLs的影響
由圖8可知,6 種AHLs(C4-HSL~C14-HSL)的混合標準品完全分離,峰形尖銳且對稱,其相應的保留時間依次為4.408、6.254、8.305、10.685、13.374、16.882 min。由于菌株的生長時間不同,所分泌的信號分子含量也有所差異,0~4 h為菌株生長的遲滯期,4~24 h進入到對數增長期。如圖9所示,在相同培養時間條件(12 h)下,熒光假單胞菌分泌量較高的信號分子為C4-HSL、C6-HSL和C8-HSL。隨富馬酸鈉質量濃度逐漸增加,各AHLs的峰面積均呈現下降趨勢,表明富馬酸鈉對熒光假單胞菌AHLs的分泌具有較好的抑制效果,推測富馬酸鈉可以作為QSI降低QS信號分子的活性。
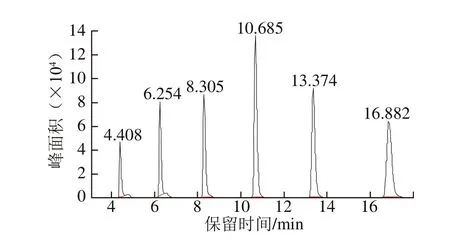
圖8 AHLs混合標準品的GC-MS圖Fig. 8 GC-MS graph of AHLs mixed with the standard
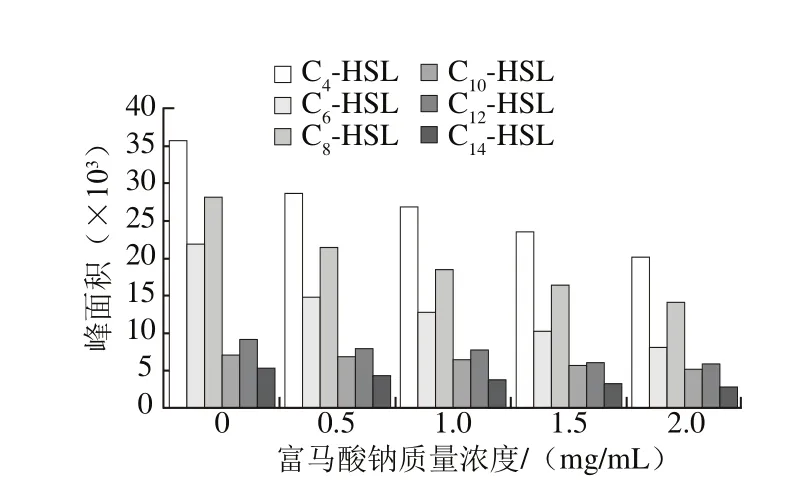
圖9 富馬酸鈉對熒光假單胞菌產AHLs的影響Fig. 9 Effect of sodium fumarate on the secretion of AHLs from Pseudomonas fluorescens
3 結 論
目前,以天然QSI或QSI類似物為靶點來干擾或阻斷QS系統已成為近年來的研究熱點。本研究以富馬酸鈉為研究對象,利用報告菌株CV026來驗證亞抑菌濃度下富馬酸鈉的QS抑制活性,探究了富馬酸鈉對熒光假單胞菌QS現象及其腐敗活性的抑制作用。結果表明,富馬酸鈉在不影響細菌生長能力條件下明顯抑制了CV026菌株紫色菌素的產生,當質量濃度為2.0 mg/mL時,與對照組相比其抑制率達50.95%;且在該質量濃度下,熒光假單胞菌生物被膜形成率降低至31.82%。因此,推測富馬酸鈉可能通過干擾熒光假單胞菌QS系統對其生物被膜、胞外蛋白酶、群集與泳動等相關腐敗特性進行抑制,且隨著質量濃度增加其抑制能力增強。此外,由GC-MS檢測結果可知,富馬酸鈉對熒光假單胞菌AHLs的分泌有較好的抑制效果。由此說明,富馬酸鈉具有較強的QS抑制活性,可作為新型的QSI用于延長水產品貨架期。本實驗主要研究了富馬酸鈉對熒光假單胞菌QS現象及腐敗特性的影響,其作用機制未明確。下一步可利用轉錄組學技術,從分子層面明晰富馬酸鈉的抑制機理。
猜你喜歡
天天愛科學(2022年9期)2022-09-15 01:12:54
天天愛科學(2022年4期)2022-05-23 12:41:48
當代水產(2022年3期)2022-04-26 14:26:56
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
科學大眾(2021年9期)2021-07-16 07:02:54
軍事文摘(2020年20期)2020-11-28 11:42:50
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
航空世界(2020年10期)2020-01-19 14:36:20
中國生殖健康(2019年2期)2019-08-23 08:12:08
產品可靠性報告(2017年7期)2017-09-05 09:49:12
