青蛤多肽的酶法制備及對前列腺癌DU-145細胞的抑制活性
2019-07-01 07:50:36張亞茹閆海強楊最素余方苗丁國芳龔戩芳
食品科學 2019年11期
關鍵詞:質量
張亞茹,閆海強,楊最素,余方苗,丁國芳,3,龔戩芳,*
(1.浙江海洋大學食品與醫藥學院,浙江省海洋生物醫用制品工程技術研究中心,浙江 舟山 316022;2.浙江海洋大學東海科學技術學院,浙江 舟山 316004;3.浙江省海洋水產研究所,浙江 舟山 316022)
前列腺癌發病率在男性惡性腫瘤發病率中位居首位,易發于老年男性。在世界范圍內,前列腺癌的發病率因環境、基因、飲食等因素不同而有很大差異[1-3]。在我國,前列腺癌發病率呈逐年上升趨勢,居泌尿系統腫瘤發病率的首位,嚴重威脅老年男性的健康[4-5]。傳統治療方法如手術、放療和冷凍治療等效果并不理想,復發率較高,并且當復發的前列腺癌轉化為非雄激素依賴性時將更難治療;而化療藥物產生耐藥性后療效更差且毒副作用明顯。因此,尋找抗前列腺癌藥物成為學者們研究的熱點。海洋是發現新型抗癌藥物的豐富的資源寶庫,其中,海洋多肽類物質因其分子質量小、活性高、毒性低等優點日益成為國內外學者研究的焦點。從雙殼貝類中提取的文蛤多肽Mere15、菲律賓蛤仔寡肽對人慢性骨髓性白血病K562細胞、前列腺癌細胞具有增殖抑制作用,并能夠濃度依賴性地誘導細胞凋亡[6-7]。
青蛤(Cyclina sinensis)屬于軟體動物門鰓瓣綱異齒亞綱簾蛤目簾蛤科,含有高含量的蛋白和不飽和脂肪酸,味道鮮美,具有很高的營養價值[8-10]。青蛤在民間具有悠久的入藥歷史,是一種重要的海洋藥物,具有軟堅散結、清熱燥濕及鎮咳的作用[11-13]。目前,已證實青蛤中富含的蛋白質、多糖和脂類具有良好的生物學活性,如抗腫瘤、抗氧化、免疫調節、抗炎等[14-18]。Jiang Changxing等[19-21]從青蛤中提取出的多糖具有抗氧化和保肝活性,對人胃癌BGC-823細胞的生長具有強烈的抑制作用。趙莎莎等[22]利用堿性蛋白酶酶解青蛤內臟獲得的水解物具有較好的體外抗氧化能力。葉盛旺等[23]發現青蛤肉經酶解法制備的青蛤多肽對RAW 264.7巨噬細胞有較好的免疫調節作用,認為其具有激活巨噬細胞、增強機體免疫力的潛在作用。酶法提取生物活性肽具有特異性強、效果好、副反應少、能耗低、易于被人體吸收等優點,并且酶法水解蛋白還因具有安全性高、價廉、易于推廣等特性而成為當前研究的熱點。然而,目前關于提取青蛤抗腫瘤活性多肽的報道并不多,本研究采用蛋白酶酶解青蛤內臟,采用正交試驗方法獲得最佳酶解條件,經分離純化篩選出活性多肽,探討其體外抗人前列腺癌DU-145細胞的活性,以期為青蛤活性多肽的制備及其抗癌作用研究提供實驗依據。
1 材料與方法
1.1 材料與試劑
青蛤購于舟山市菜市場,經浙江海洋大學趙盛龍教授鑒定為青蛤。人前列腺癌DU-145細胞購于中國科學院上海生命科學院生化細胞所,由浙江省海洋生物醫用制品工程技術研究中心傳代保存。
堿性蛋白酶、胰蛋白酶、木瓜蛋白酶、中性蛋白酶、胃蛋白酶 北京亞太恒信生物科技有限公司;F12培養基 美國Gibco公司;胎牛血清 杭州四季青生物工程公司;噻唑藍(3-(4,5-dimethylthiazol-2-yl)-2,5-diphenyltetrazolium bromide,MTT)、胰蛋白酶美國Sigma公司;吖啶橙/溴化乙錠(acridine orange/ethidium bromide,AO/EB) 杭州昊天生物技術有限公司;JC-1細胞線粒體膜電位試劑盒 上海貝博生物公司;非轉移性克隆23型(nm23H1)單克隆抗體(工作濃度按體積比1:100稀釋) 美國Santa Cruz Biotechnology公司;免疫組織化學試劑盒 丹麥DAKO有限公司;其他試劑均為國產分析純。
1.2 儀器與設備
CF16RXⅡ型高速低溫離心機、H-7650透射電子顯微鏡 日本日立公司;Forma3111 CO2培養箱 美國Thermo Scientific公司;Cogent u Scale超濾系統 德國默克密理博公司;Apurifier UPC 100快速蛋白液相色譜系統 GE醫療生命科學有限公司;1260高效液相色譜(high performance liquid chromatography,HPLC)儀美國安捷倫公司;超凈工作臺 上海智城分析儀器制造有限公司;CKX4倒置顯微鏡、BX2-FLB3熒光顯微鏡、CCD-NC6051顯微攝像儀 日本Olympus公司;酶標儀美國Bio-Rad公司;easy Cyte6 HT-2L流式細胞儀 美國Millipore公司。
1.3 方法
1.3.1 細胞培養
將DU-145細胞培養在含雙抗(100 U/mL青霉素、100 U/mL鏈霉素)和質量分數10%胎牛血清的F12培養液中,放置在37 ℃、含有體積分數5% CO2的培養箱中孵育,當細胞密度達到80%以上時用質量分數0.25%胰蛋白酶消化傳代,取對數生長期的細胞進行實驗。
1.3.2 青蛤內臟酶解液的制備
1.3.2.1 最佳酶種類的選擇
分別取10.0 g勻漿青蛤內臟,以堿性蛋白酶、胰蛋白酶、木瓜蛋白酶、中性蛋白酶和胃蛋白酶這5 種酶為供試酶,在各自最適酶解溫度及pH值條件(表1)下,加1 200 U/g、保溫6 h進行酶解。隨后滅活15 min,于4 ℃、12 000 r/min離心10 min,取其上清液,采用甲醛電位滴定法測定氨基氮(amino nitrogen,ANN)含量。
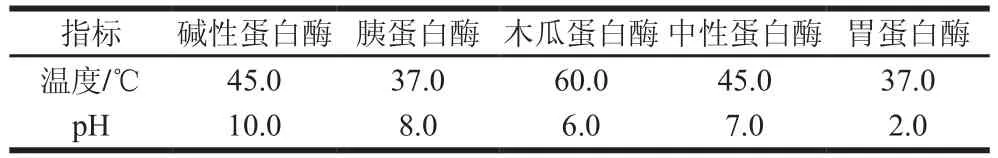
表1 5 種蛋白酶的最適酶解溫度和pH值Table 1 Optimum temperature and pH of proteases
1.3.2.2 酶解條件的優化
在初步確定酶種類的基礎上,以對DU-145細胞的增殖抑制率(IR)為指標,以料液比、pH值、加酶量、溫度、時間5 個因素進行L16(45)正交試驗(表2),確定最優酶解條件。
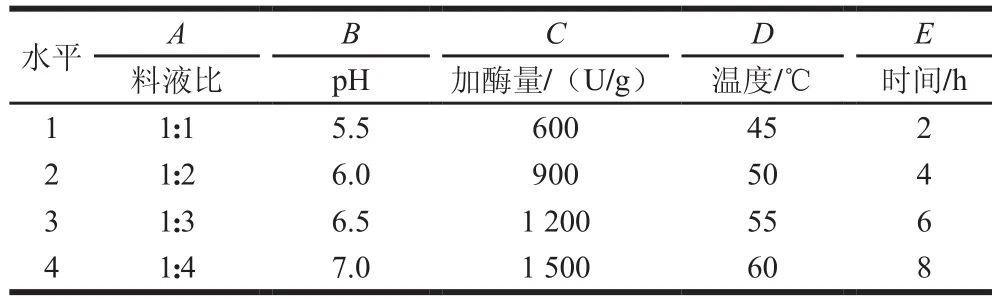
表2 木瓜蛋白酶酶解青蛤內臟正交試驗因素水平Table 2 Coded levels and corresponding actual levels of independent variables used for orthogonal array design
1.3.3 酶解多肽的分離純化
1.3.3.1 超濾
取最佳酶及最優酶解條件下的上清液,用截留分子質量為8 kDa的超濾膜在Cogent u Scale超濾系統上進行超濾,得到小于8 kDa和大于8 kDa的兩種超濾液。再用截留分子質量為5 kDa的超濾膜對小于8 kDa的部分超濾,截留獲得大于5 kDa且小于8 kDa和小于5 kDa的兩種超濾液。換用截留分子質量為3 kDa的超濾膜對小于5 kDa的部分超濾,截留獲得大于3 kDa且小于5 kDa和小于3 kDa的兩種超濾液。分別取不同分子段酶解液冷凍干燥樣品配制成質量濃度15 mg/mL進行MTT實驗,篩選出對DU-145細胞抑制率最高的超濾組分備用。
1.3.3.2 瓊脂糖凝膠層析
取1.3.3.1節的超濾組分進行瓊脂糖凝膠層析分離,酶解液質量濃度為0.05 g/mL,離心后取上清液過0.22 μm濾膜,在Apurifier UPC 100快速蛋白色譜系統上進行洗脫。色譜柱規格:300 mm×10 mm;色譜柱填料:瓊脂糖凝膠;色譜柱料顆粒尺寸:(10±2)μm;上樣量:500 μL;流動相:超純水;洗脫液流速:0.5 mL/min;檢測波長:280 nm;自動收集體積:3.2 mL/管。收集各峰溶液進行冷凍干燥,采用MTT法檢測抑制率最高的峰組分。
1.3.3.3 HPLC法分離制備及純度檢測
取1.3.3.2節抑制率最高的峰組分通過HPLC進一步分離,其質量濃度為1.0 mg/mL。色譜柱:ZORBAX SB-C18分析型色譜柱(250 mm×4.6 mm,5 μm);檢測波長:214、280 nm;流速:0.8 mL/min;流動相A為乙腈(含0.05%三氟乙酸),流動相B為超純水(含0.06%三氟乙酸),采用梯度洗脫方式:0%(體積分數,下同)流動相B,4 min;0%~100%流動相B,25 min;100%流動相B,6 min,柱溫:25 ℃;自動進樣,進樣量:100 μL。收集MTT法檢測抑制率最高的峰組分,冷凍干燥,即得到青蛤多肽(Cyclina sinensis peptides,CSP)。
1.3.4 DU-145細胞抑制活性檢測
1.3.4.1 細胞增殖抑制實驗
將DU-145單細胞懸液以1×104個/mL的密度每孔200 μL接種至96 孔板。24 h后棄去培養液,設置空白對照組(正常培養基培養)及加藥組(CSP質量濃度分別為2、4、8、12、15 mg/mL),每組設5 個平行孔。培養24 h后去培養液,每孔加含質量分數10% MTT的磷酸鹽緩沖液(phosphate buffered saline,PBS)200 μL,繼續孵育4 h。實驗結束后棄去孔中液體,每孔加二甲基亞砜150 μL,充分振蕩10 min。采用酶標儀于490 nm波長處測定各孔的OD值,每組實驗重復3 次。按下式計算細胞增殖抑制率并計算其半數抑制濃度(half maximal inhibitory concentration,IC50)。

1.3.4.2 倒置顯微鏡觀察細胞形態
將經泡酸、消毒處理后的蓋玻片置于6 孔板內,接種密度為1×105個/mL的DU-145細胞懸液,常規孵育24 h,棄去孔中液體,設空白對照組(正常培養基培養)和加藥組(CSP質量濃度分別為2、8、12 mg/mL),培養24 h,于倒置顯微鏡下觀察各組細胞形態變化。
1.3.4.3 Hoechst 33258熒光染色觀察
細胞接種密度、實驗分組及藥物處理同1.3.4.2節方法。24 h后終止培養,棄去各孔中液體并用體積分數2.5%戊二醛固定20 min,pH 7.2 PBS洗2 次,0.5 μg/mL Hoechst 33258熒光染液室溫染色15 min,然后用PBS洗2 次,于熒光顯微鏡下觀察拍照。
1.3.4.4 ROS檢測
活性氧(reactive oxygen species,ROS)檢測中細胞接種密度、實驗分組及藥物處理同1.3.4.2節方法。培養24 h后,細胞用胰酶消化并進行離心收集(1 000 r/min、6 min),按照體積比1:1 000用無血清的F12營養液和2,7-二氯二氫熒光素二乙酸酯(2,7-dichlorodihydrofluorescein diacetate,DCFH-DA)均勻懸浮細胞,濃度約為2×106個/mL。于37 ℃、5% CO2細胞培養箱內孵育20 min,為了使探針和細胞更充分地接觸,每隔3~5 min將離心管上下顛倒混勻。孵育結束后用不含有血清的F12營養液洗滌細胞3 次,用來充分除去沒有進入細胞中的DCFH-DA。于熒光顯微鏡下觀察并拍照。
1.3.4.5 透射電子顯微鏡觀察細胞微觀形態
DU-145細胞接種于培養瓶中,設空白對照組(正常培養基培養)和加藥組(CSP質量濃度分別為2、8 mg/mL)。培養24 h,取消化后的細胞懸液以體積分數2.5%戊二醛及鋨酸雙重固定,丙酮梯度脫水,中性618樹脂包埋。超薄切片機切片,于透射電子顯微鏡下觀察并拍照。
1.3.4.6 流式細胞儀檢測細胞線粒體膜電位變化
細胞接種密度、實驗分組及藥物處理同1.3.4.2節方法。取消化的細胞懸液,于2 000 r/min離心5 min,PBS洗2 次。按試劑盒說明書操作,取1 μL JC-1試劑加入500 μL 1×Incubation Buffer混勻,10 000 r/min離心1 min,上清液即JC-1工作液;取500 μL JC-1工作液懸浮細胞并孵育20 min,室溫下2 000 r/min離心5 min,并用1×Incubation Buffer洗細胞,重復2 次。混勻懸浮細胞并過200 目篩,于流式細胞儀進行檢測。
1.3.4.7 免疫組織化學法檢測DU-145細胞nm23H1蛋白的表達
細胞接種密度、實驗分組及藥物處理同1.3.4.2節方法。取出蓋玻片,用丙酮固定20 min。加入體積比1:100的單克隆抗體4 ℃過夜;滴加EnVision液室溫孵育30 min;使用二氨基聯苯胺進行顯色,光學顯微鏡下觀察控制顯色時間,自來水沖洗;蘇木精復染5 min后用乙醇梯度脫水、二甲苯透明,并用中性樹膠封片。于光學顯微鏡下觀察并拍照。
1.4 數據統計與分析
實驗結果以 ±s表示,數據使用SPSS 18.0軟件進行單因素方差分,P<0.05表示差異有顯著性。采用Excel軟件作圖。
2 結果與分析
2.1 酶解條件的優化
2.1.1 5 種蛋白酶的酶解結果
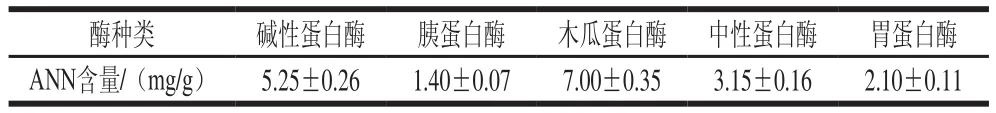
表3 5 種蛋白酶酶解青蛤內臟的ANN含量Table 3 Comparison of amino nitrogen contents of Cyclina sinensis hydrolysates produced using five proteases
每種酶在其最適的酶解溫度、pH值條件下可以得到最好的酶解效果,即以最少的投料使酶解產物含量最大化,使水解徹底。ANN含量與水解度成正相關,在各自最適酶解條件下,如表3所示,木瓜蛋白酶酶解物的ANN含量最高,達到了(7.00±0.35)mg/g,堿性蛋白酶次之。說明木瓜蛋白酶對青蛤內臟的水解效果最好,故選定木瓜蛋白酶為本實驗最佳酶。
2.1.2 木瓜蛋白酶酶解青蛤內臟的正交試驗結果
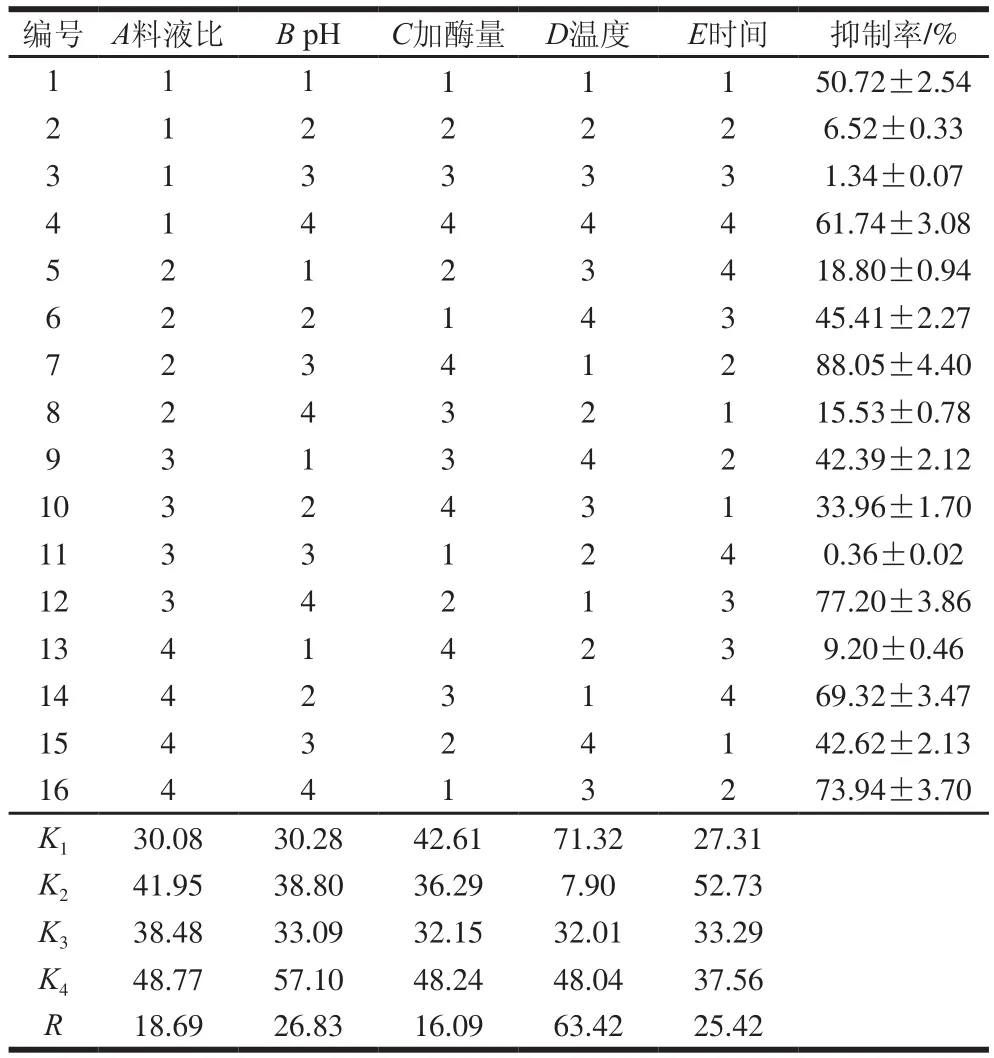
表4 木瓜蛋白酶酶解青蛤內臟正交試驗結果Table 4 Orthogonal array design with experimental results
如表4所示,從抑制率大小可知,5 個因素對酶解效果皆有影響,由極差(R)大小得D(溫度)>B(pH值)>E(時間)>A(料液比)>C(加酶量),且A4B4C4D1E2組合效果最好,即料液比1:4、pH 7.0、加酶量1 500 U/g、溫度45.0 ℃、酶解時間4 h。每個實驗重復3 次,所得結果均為此組合效果最佳,即正交試驗結果具有可靠性。
2.2 酶解液的分離純化結果
2.2.1 超濾截留結果
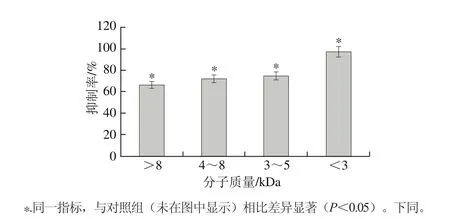
圖1 酶解液的超濾截留組分對DU-145細胞的增殖抑制作用Fig. 1 Inhibitory effect of hydrolysate fractions obtained by ultrafiltration on DU-145 cells
如圖1所示,小于3 kDa的酶解液對DU-145細胞的抑制率最高,達到(89.46±4.47)%。故選定小于3 kDa酶解液進行下步分離純化。
2.2.2 Superose 12 10/300 GL層析分離純化結果
取小于3 kDa冷凍干燥樣品進行瓊脂糖凝膠層析,于280 nm波長處出現3 個峰,即峰1、峰2和峰3,如圖2所示。MTT法顯示峰1對DU-145細胞的抑制活性最高。
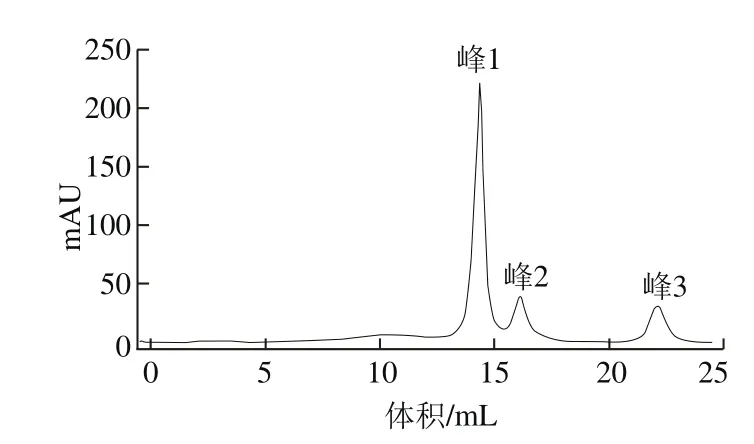
圖2 瓊脂糖凝膠柱洗脫峰Fig. 2 Elution peaks on agarose gel column
2.2.3 HPLC法分離制備及純度檢測結果
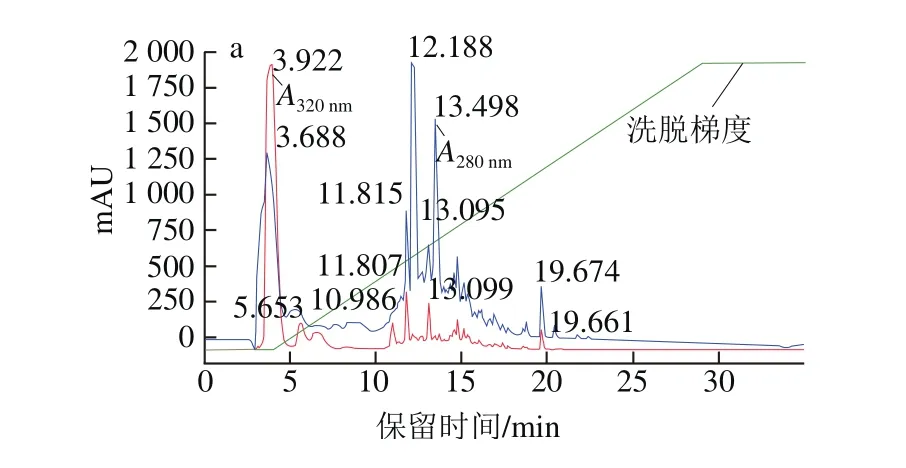
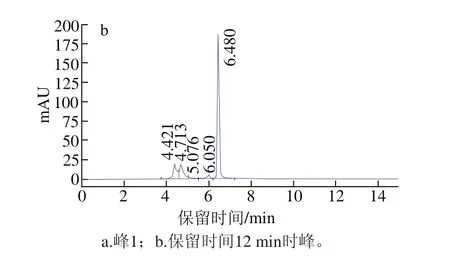
圖3 峰1組分及保留時間12 min時峰組分的高效液相色譜圖Fig. 3 High performance liquid chromatography of peak 1 and the peak of retention time 12 min
選取圖3a中保留時間約為12 min的峰組分(峰面積16.32,峰高1 999.79)進行收集,經N端測序得出該肽的氨基酸序列為:Ile-Leu-Tyr-Met-Pro,分子質量635.82 Da,命名為CSP。收集12 min時峰純品再次采用HPLC法檢測純度,在280 nm波長處約7 min時出現主要單一峰,如圖3b所示。
2.3 細胞增殖抑制實驗結果

圖4 CSP對DU-145細胞的增殖抑制作用Fig. 4 Antiproliferative effect of CSP on DU-145 cells
如圖4所示,隨CSP質量濃度的增加和作用時間的延長對DU-145細胞的增殖抑制率明顯上升。當CSP質量濃度為2 mg/mL、作用時間為24 h時抑制率為(9.89±0.49)%;而當CSP質量濃度為15 mg/mL、作用時間72 h時抑制率達到(84.17±4.21)%,IC50為(5.36±0.27)mg/mL,與對照組相比差異顯著(P<0.05)。
2.4 倒置顯微鏡觀察細胞形態結果
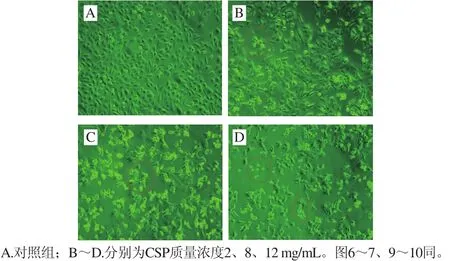
圖5 倒置顯微鏡下DU-145細胞的形態變化(×200)Fig. 5 Morphologic changes of DU-145 cells under inverted microscope (× 200)
如圖5所示,對照組DU-145細胞形態飽滿、生長良好,視野下細胞數目較多(圖5A);2 mg/mL組細胞間隙增大、輪廓模糊(圖5B);而高質量濃度(12 mg/mL)組細胞變圓、變亮,形態更加不規則,細胞數目明顯少于對照組,同時可見較多懸浮在培養液中的死細胞(圖5D)。
2.5 Hoechst 33258熒光染色結果
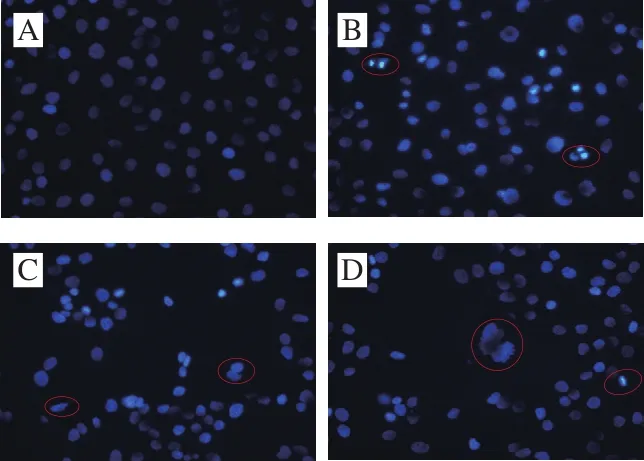
圖6 Hoechst 33258染色觀察DU-145細胞形態變化(×400)Fig. 6 Morphologic changes of DU-145 cells observed by Hoechst 33258 staining (× 400)
如圖6所示,對照組細胞核呈現出淡藍色熒光,且細胞形態規則;而CSP作用后可見個別細胞核皺縮呈亮藍色;隨著質量濃度的增加,視野中細胞數減少,部分核呈碎片狀或點狀且形態不規則,藍色熒光增加,同時出現了凋亡小體。
2.6 ROS檢測結果

圖7 CSP誘導DU-145細胞中ROS的產生Fig. 7 CSP induces ROS generation in DU-145 cells
如圖7所示,對照組出現極少量的綠色熒光;與對照組比較,經CSP作用后熒光強度明顯增加,且隨著質量濃度的增加熒光強度依次增強。這表明DU-145細胞內ROS的產生量隨著CSP質量濃度的增加呈明顯增加趨勢。
2.7 透射電子顯微鏡觀察細胞微觀形態結果
如圖8所示,對照組DU-145細胞培養24 h后可見細胞表面有微絨毛,胞核形態規則,具有完整的核膜,染色質均勻分布,含有一個核仁。CSP處理后細胞的超微結構發生明顯改變,染色質電子密度增高并濃縮聚集至核膜邊緣,部分細胞膜有“出芽”現象,出現凋亡小體;隨著CSP質量濃度的增加,細胞的微絨毛消失,細胞質內可見大量空泡形成,并出現了大小不一的自噬體,細胞核固縮明顯,表現出凋亡細胞的特點。
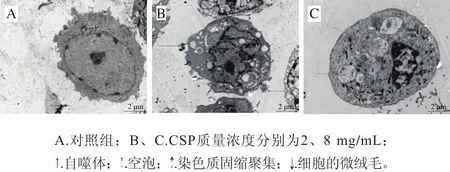
圖8 CSP作用24 h后DU-145細胞的超微結構(× 3 700)Fig. 8 Ultrastructural changes of DU-145 cells incubated with CSP for 24 h (× 3 700)
2.8 細胞線粒體膜電位變化結果
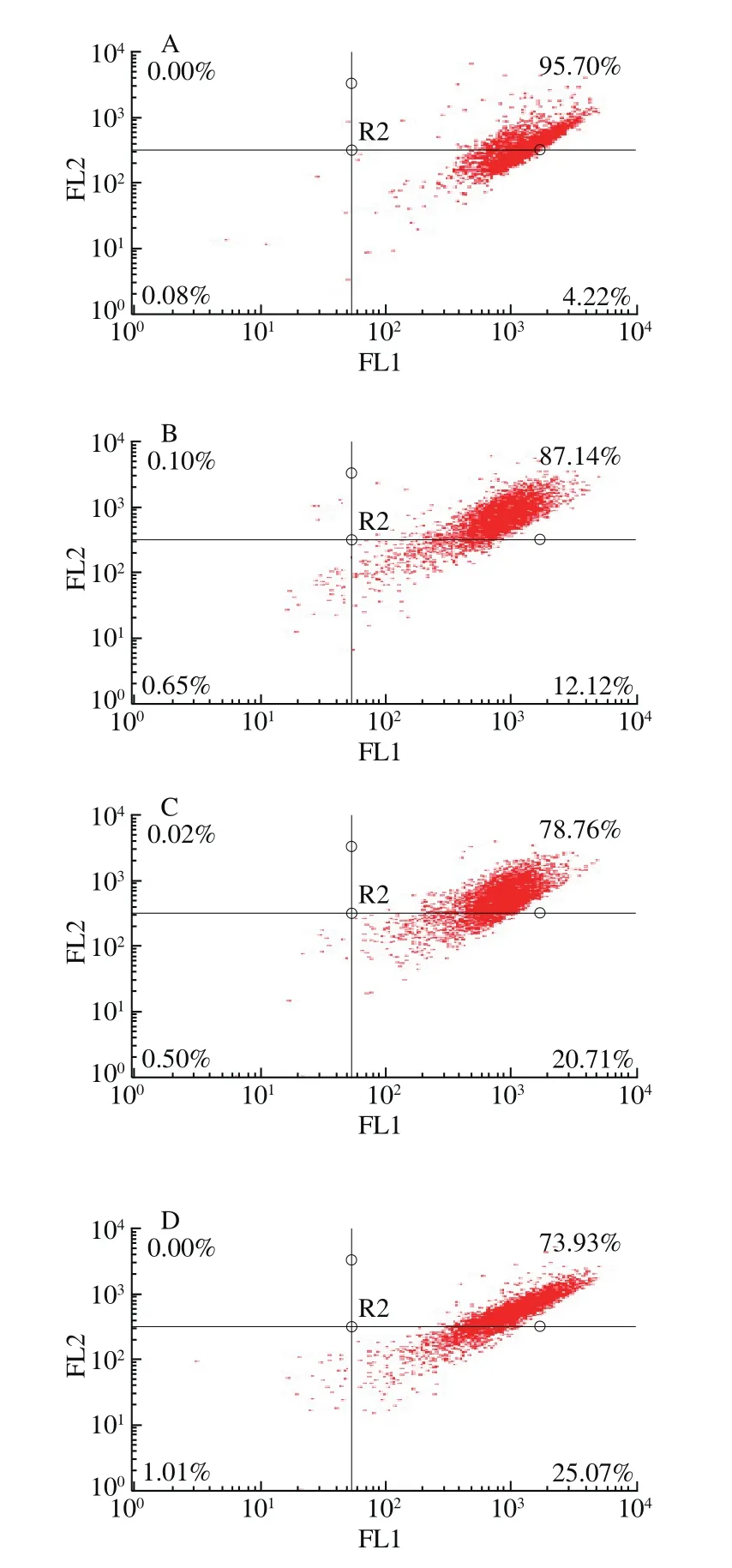
圖9 CSP對DU-145細胞線粒體膜電位的影響Fig. 9 Changes in mitochondrial membrane potential of DU-145 cells treated with CSP
從圖9可以看出,右上象限表示正常的膜電位比例,右下象限表示下降的膜電位比例。與對照組相比,2、8 mg/mL和12 mg/mL組膜電位下降率分別為12.12%、20.71%和25.07%。隨CSP質量濃度的增大,線粒體膜電位下降率隨之增大,即發生細胞凋亡的細胞數呈顯著性增加趨勢。
2.9 免疫組織化學法檢測CSP對DU-145細胞nm23H1蛋白的陽性表達結果

圖10 CSP對DU-145細胞nm23H1蛋白表達的影響(×400)Fig. 10 Changes in expression of nm23H1 in DU-145 cells treated with CSP (× 400)
如圖10所示,nm23H1蛋白陽性表達為細胞質內出現棕色結構對照組大部分細胞胞漿內呈淺棕色,個別細胞呈現陰性(圖10A)。隨著CSP質量濃度的增加,視野下的細胞數量明顯減少,細胞質內的陽性部位顏色逐漸加深,出現了巨核細胞,部分細胞核呈固縮狀態。
3 討 論
近年來研究發現海洋生物活性肽具有較好的抗腫瘤作用[24-26],其體外抗腫瘤細胞活性常用MTT法進行篩選[27]。本研究采用酶解法提取CSP,經MTT法檢測,結果顯示,隨著CSP質量濃度的增加和作用時間的延長,其對DU-145細胞的增殖抑制率明顯上升,且具有明顯的時間及劑量依賴性。當CSP質量濃度為15 mg/mL、作用72 h后,抑制率高達84.17%,IC50為5.36 mg/mL。同時DU-145細胞的形態也發生變化,表現為細胞間隙變大、形態不規則、核固縮、細胞質內出現空泡和形成凋亡小體,其中凋亡小體的形成表明CSP抑制DU-145細胞的增殖與細胞凋亡相關。
ROS是誘導細胞發生凋亡的一種重要因素,因其通過與細胞內的脂質、核酸、蛋白質發生反應,導致機體組織脂質過氧化、DNA氧化損傷和細胞內蛋白變性,對細胞造成損傷;ROS還可作為細胞內信使,活化許多信號傳導通路如細胞凋亡通路,間接導致細胞損傷[28],從而使細胞內產生更多ROS。在ROS積聚的情況下,除了主動的細胞凋亡外,其能通過激活一系列信號通路來誘導細胞凋亡,導致細胞凋亡失調而使得細胞過度凋亡,引發自身免疫疾病和炎癥。本研究結果表明,CSP作用后DU-145細胞ROS的產生量明顯增加,且呈劑量依賴性。研究發現藥物抗腫瘤作用的機制與其作用于線粒體導致功能紊亂、抑制腫瘤細胞血管生成能力等相關[29],而線粒體膜電位(線粒體膜通透性)的降低是細胞凋亡早期的不可逆事件。經流式細胞術檢測,與對照組細胞相比,CSP作用后DU-145細胞的線粒體膜電位呈顯著性下降趨勢,隨著CSP質量濃度增加,其膜電位下降率明顯增加,從4.22%增加到25.07%。因此推測CSP可能通過刺激DU-145細胞產生大量的ROS引起線粒體膜通透性改變,從而啟動細胞凋亡。由此可見線粒體凋亡的信號通路在CSP所誘導的DU-145細胞凋亡過程中發揮著關鍵性作用。
據報道,90%腫瘤病人最終的死亡原因都是腫瘤的復發、侵襲和轉移,因此防止腫瘤轉移的方法或藥物越來越受腫瘤臨床醫生及患者的重視,一些與腫瘤細胞發病機制密切相關的分子靶點不斷被發現,靶向藥物的篩選成為當前抗腫瘤藥物研究最為活躍的領域之一。nm23H1是目前研究較多的轉移抑制基因,其表達程度可作為判斷腫瘤有無轉移的重要指標。前期的研究結果顯示,nm23H1蛋白的表達與前列腺癌的發生、轉移和生存率密切相關,且與前列腺癌患者的存活率呈正相關[30]。免疫組織化學法結果表明,對照組DU-145細胞中nm23H1蛋白的表達程度較低,CSP作用后表達量隨著CSP質量濃度的增加逐漸增高。說明CSP能夠促進nm23H1蛋白在DU-145細胞中表達,對于抑制其轉移具有重要的作用,nm23H1蛋白表達量可以成為篩選CSP活性的指標。
4 結 論
本實驗采用木瓜蛋白酶,在料液比1:4、pH 7.0、加酶量1 500 U/g、溫度45.0 ℃、酶解時間4 h條件下對青蛤內臟勻漿液進行酶解,經超濾截留、瓊脂糖凝膠層析、HPLC分離純化最終獲得CSP,經氨基酸測序得到其氨基酸序列為Ile-Leu-Tyr-Met-Pro。研究結果表明,CSP能抑制DU-145細胞的增殖,且呈現時間與劑量依賴,并使細胞出現凋亡的形態學特征,其作用機制與DU-145細胞ROS、nm23H1蛋白表達量的增加以及線粒體膜電位的降低相關。至于其通過何種信號傳導通路引起凋亡的機制有待于后續研究證實。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
