飼料脂肪源對大菱鲆生長、體組成及血清生化指標的影響
2019-07-12 07:59:40趙曼茜張斐然吳立新李思萌李笑迎
中國飼料 2019年13期
趙曼茜, 張斐然, 吳立新*, ,李思萌 ,2, 李笑迎
(1.大連海洋大學遼寧省水生生物學重點實驗室,遼寧大連116023;2.聊城市開發區實驗中學,山東聊城 252000)
飼料中的脂肪是魚類必需脂肪酸的來源,具有多種生理功能,是魚類重要的營養成分之一。有研究表明,不同脂肪源對魚類生長、體組成和血清生化指標影響不同。馮健和覃志彪(2006)的試驗結果表明,太平洋鮭(Oncorhynchus spp.)日糧中脂肪以添加大豆磷脂的生長性能最好;陳家林等(2011)研究發現,豆油、椰子油和菜籽油是異育銀鯽 (Carassius auratus gibelio)飼料中良好的脂肪源;吉紅等 (2009)建議生產中使用豆油作為鯽(Carassius auratus)飼料的脂肪源等。因此,研究不同魚類適宜的脂肪源意義重大。
大菱鲆(Scophthalmus maximus)又名多寶魚,是我國北方最重要的海水養殖魚類之一 (馬愛軍等,2001)。本試驗以魚油(FO)、豆油(SO)、菜籽油(RO)、花生油(PO)、豬油(L)、魚油-豆油(FSO)(1∶1)和混合油(MIX)(魚油∶豆油∶豬油=3∶4∶3)為脂肪源的7種飼料,研究不同脂肪源對大菱鲆生長、體組成以及血清生化指標的影響,為篩選合適的大菱鲆飼料脂肪源提供一定的理論依據。
1 材料與方法
1.1 試驗飼料的配制 本試驗共配制了7種飼料,除脂肪源不同外,其余飼料成分相同,試驗飼料組成及營養水平見表1。飼料蛋白源主要由魚粉、豆粕提供,分別添加 6%的魚油(FO)、豆油(SO)、菜籽油(RO)、花生油(PO)、豬油(L)、魚油-豆油 (1:1)(FSO) 和魚油-豆油-豬油(3:4:3)(MIX)。飼料原料粉碎后過60目篩,按試驗配比充分混勻,用制粒機制作成粒徑為2 mm的顆粒飼料,自然風干后存放于-20℃冰箱備用。
1.2 試驗魚與飼養管理 試驗用大菱鲆購自大連天正實業有限公司。以基礎飼料(FO)馴養兩周。試驗開始前,將魚饑餓24 h,選取體質健康、規格均勻的魚[(27.31±0.10)g]252 尾。試驗共設 7組,每組設3個重復,共21個水族箱(60 cm×45 cm×40 cm,實際水量90 L),每個水族箱放入12尾魚,飼養40 d。試驗期間,以砂濾后的海水為水源,日換水一次,換水量為1/3~1/2。水溫控制在14.0~ 19.0℃,溶解氧>6.0 mg/L,氨氮<0.1 mg/L,pH 8.0~8.1,光照采用日光燈控制,周期為12 L/12 D。每日飽食投喂兩次(07:30和17:30),投喂后30 min左右收集殘餌。
1.3 樣品采集 試驗結束時將魚饑餓24 h,將每箱魚分別稱重(精確至0.01 g)并隨機取3尾裝于自封袋,置于-20℃冰箱中保存,用于常規體成分測定。每箱隨機另取2尾于冰盤解剖,取其背肌、內臟和空殼,裝入對應自封袋中置于-20℃冰箱待測。每箱隨機另取2尾尾靜脈取血,經3500 r/min離心15 min制得血清,置于-20℃冰箱待測。
1.4 樣品分析測定 飼料和魚體常規成分測定參照AOAC(1995)。水分通過在105℃烘箱烘至恒定質量測定;粗蛋白質含量用凱氏定氮儀測定(總氮×6.25);粗脂肪含量采用索氏抽提法,以乙醚為抽提液測定;粗灰分含量通過在馬福爐550℃灼燒(8 h)測定;血清生化指標由大連市體檢中心測定。每個樣品重復測定2次,如果相對偏差超過2%,則增加重復測定次數,采用兩個相對偏差在2%以下的測定值平均數作為測定結果。
1.5 計算公式
特定生長率/(%/d)=(lnWt-lnW0)/t×100;
蛋白質效率/%=(Wt-W0)/(Wd×Cp)×100;
肥滿度/%=Wt/L3×100;
飼料系數=Wd/(Wt-W0)。
式中:W0和Wt分別為試驗開始和結束時魚的濕重,g;Wd為攝入的飼料干重,g;Cp為飼料蛋白質含量,%;L 為魚體長,cm;t為試驗天數,d。
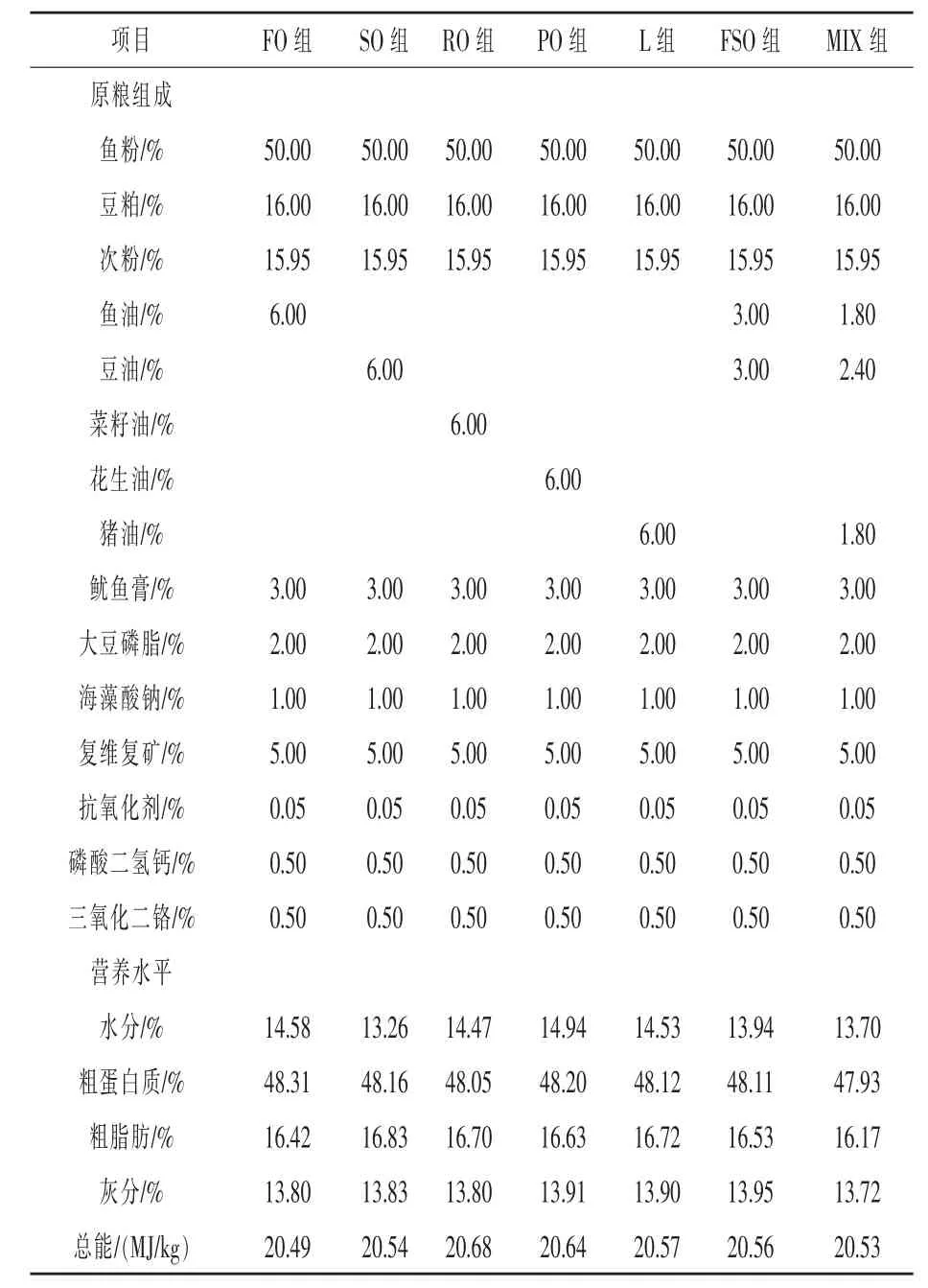
表1 試驗飼料組成及營養水平(風干基礎)
1.6 統計分析 采用SPSS 17.0統計軟件對所得數據進行單因素方差分析(one-way ANOVA),若差異顯著(P<0.05),則進行Duncan's多重比較檢驗組間的差異,統計結果以“平均值±標準誤”表示。
2 結果
2.1 生長性能 如表2所示,魚油-豆油組的特定生長率顯著高于其他各組,較魚油組(基礎飼料組)提高了10.42%(P<0.05),豬油組特定生長率最低,較魚油組降低了19.79%(P<0.05);從蛋白質效率來看,除了魚油-豆油組顯著高于混合油組外 (P<0.05),其他各組間無顯著差異 (P>0.05);飼料系數以魚油-豆油組最低,但與魚油、豆油、菜籽油組之間無顯著差異(P>0.05),豬油、混合油、花生油組飼料系數分別較魚油組提高了36.36%、33.33%、25.25%(P < 0.05)。
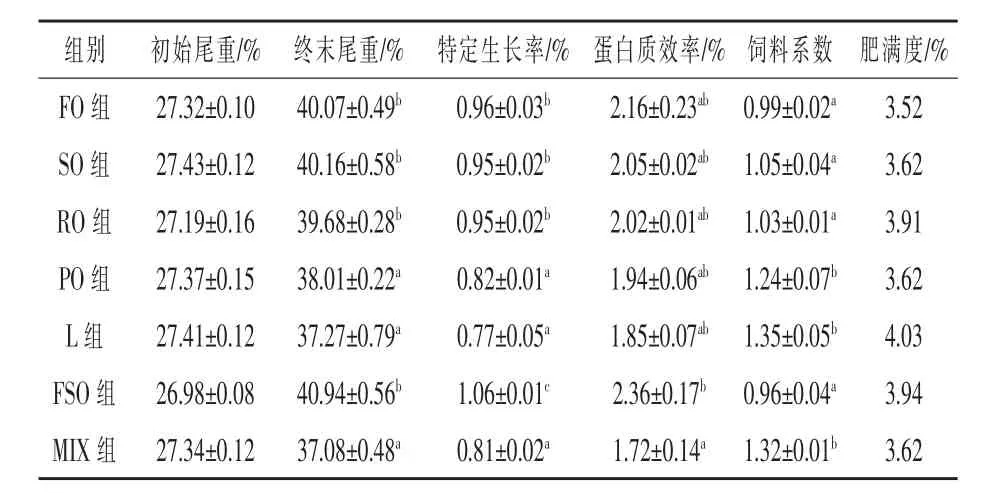
表2 飼料脂肪源對大菱鲆生長和飼料利用的影響
2.2 體組成和體組織水分、脂肪含量 如表3所示,飼料脂肪源對魚體水分、粗脂肪和粗灰分含量均沒有顯著影響(P>0.05)。豬油組粗蛋白質含量顯著低于菜籽油組、花生油組和魚油組,其中較魚油組降低了4.47%(P<0.05),其余各組間無顯著差異(P>0.05)。從表4可以看出,除豬油組魚體空殼的水分顯著高于菜籽油組和混合油組 (P<0.05)外,飼料脂肪源對大菱鲆空殼、內臟、背肌的水分和脂肪含量沒有顯著影響(P>0.05)。
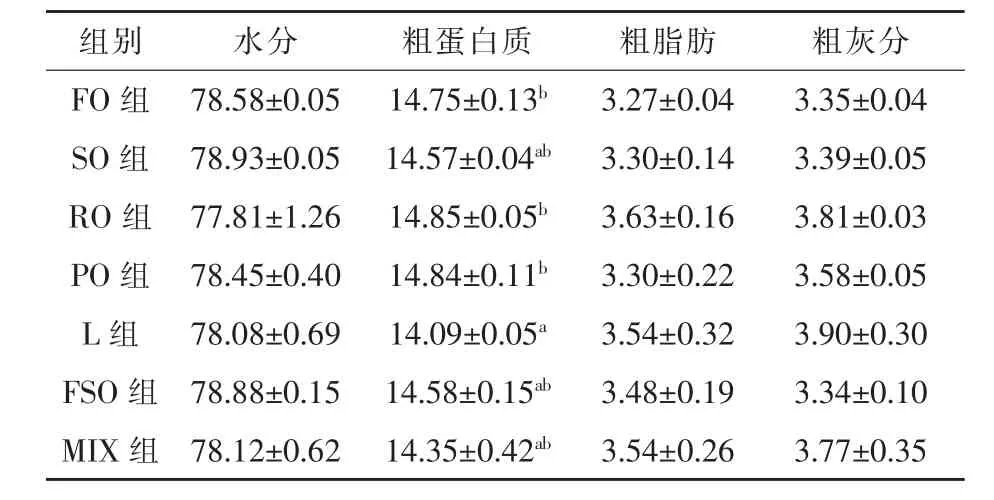
表3 飼料脂肪源對大菱鲆魚體生化組成的影響%濕重
2.3 血清生化指標 如表5所示,各組間總膽固醇、甘油三酯、高密度脂蛋白膽固醇和堿性磷酸酶無顯著差異(P>0.05)。豬油組和混合油組谷丙轉氨酶含量顯著高于其余各組,較魚油組分別升高了124.89%和119.94%(P<0.05),其余各組間沒有顯著差異(P>0.05);混合油組谷草轉氨酶含量顯著高于其余各組,較魚油組上升了352.51%(P<0.05),其余各組間無顯著差異(P>0.05)。
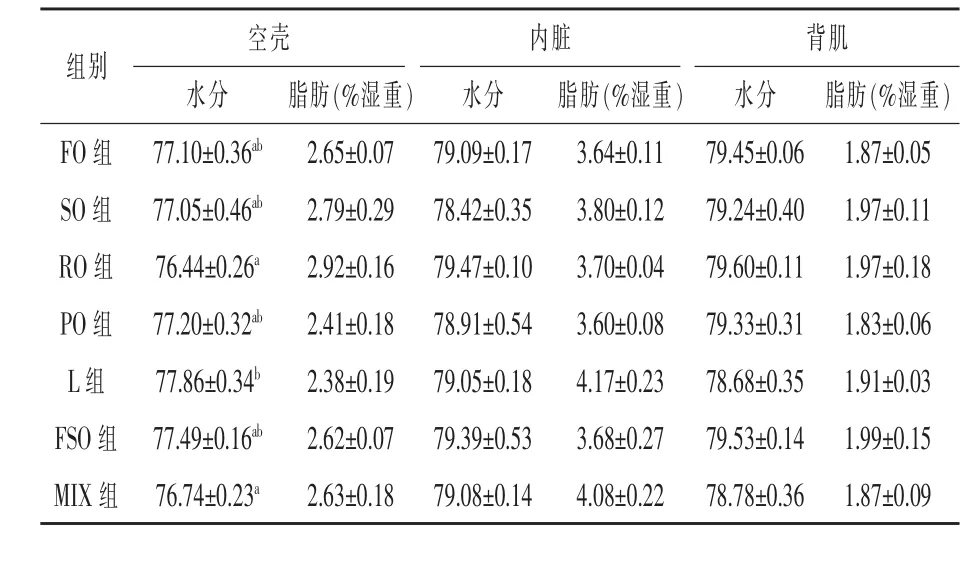
表4 飼料脂肪源對大菱鲆各組織水分和脂肪含量的影響
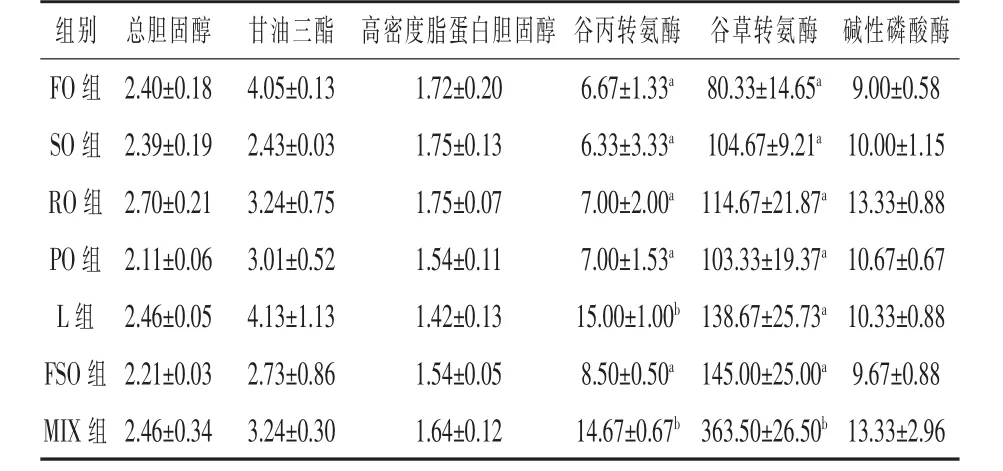
表5 飼料脂肪源對大菱鲆血清生化指標的影響U/L
3 討論
本試驗結果顯示,各組間特定生長率存在顯著差異(P<0.05),以魚油-豆油組最高,顯著高于魚油、豆油、菜籽油組和花生油、混合油、豬油組(P<0.05),這與張媛媛等(2012)用不同脂肪源飼料飼喂異育銀鯽的研究結果相似。蛋白質效率除混合油組顯著低于魚油-豆油組(P<0.05)外,其他各組間飼料營養價值與魚油無顯著差異 (P>0.05)。從飼料系數來看,大菱鲆對魚油-豆油組、魚油組、菜籽油組和豆油組飼料利用率高。不同的飼料脂肪源對魚體的肥滿度沒有顯著影響,與於葉兵等(2012)用不同脂肪源飼料飼喂異育銀鯽、杜震宇等(2002)對鱸魚(Lateolabrax japonicus)的研究結果一致。
本試驗中魚體水分、粗脂肪含量在各組間沒有顯著差異 (P>0.05),這與真鯛(Pagrosomus major)(Glencross等,2003)、 異育銀鯽 (陳家林等,2011)等的研究結果一致。通常不同脂肪源對魚體組織的脂肪含量有一定的影響(Sargent等,2002),如衛曉怡和陳舜勝(2003)研究表明,大豆油添加組團頭魴(Megalobrama amblycephala)魚體背部肌肉脂質積累較好,但在本試驗中,各試驗組空殼、內臟、背肌的脂肪含量均無顯著差異(P > 0.05)。
魚類血清生化指標能夠反映魚類對疾病的生理應答(Hrubec等,2008)。血脂含量占全身脂肪含量的很小一部分,但其代謝非常活躍,魚類腸道吸收的外源性食物脂類,肝臟合成的內源性脂類和脂肪組織的利用都要經過血液再到其他組織,脂肪的運輸也要經過血清,因此,血脂水平可以反映全身脂類代謝情況 (Hiraoka等,1979;Nakagawa,1978)。血清中總膽固醇和甘油三酯含量過高可能會導致心血管疾病,而高密度脂蛋白膽固醇可以將膽固醇從動脈壁運送到肝臟,通過肝臟轉化代謝。因此高密度脂蛋白膽固醇有減輕血液中過量的膽固醇,減慢血管炎癥,降低心血管疾病發生率的功能(孫明堂等,1982)。本試驗中,各組間的總膽固醇、甘油三酯和高密度脂蛋白膽固醇含量沒有顯著差異(P>0.05),說明這7種脂肪源不會對大菱鲆幼魚的脂肪代謝產生不利影響。谷丙轉氨酶和谷草轉氨酶存在于肝組織中,魚類血液中的谷丙轉氨酶和谷草轉氨酶活性變化可以反映肝細胞受損情況 (康格菲,1989)。在正常情況下,魚類血清中轉氨酶的活性較低,血液中只含有少量的由魚體組織細胞釋放的轉氨酶,當受損組織范圍較大或組織病變引起細胞膜通透性增加時,使細胞主動轉運體系受到損傷,導致物質主動轉運機制和滲透壓調節能力削弱從而消失,甚至細胞解體,血液中滲入由組織細胞內釋放的大量谷丙轉氨酶和谷草轉氨酶,導致血液中谷丙轉氨酶和谷草轉氨酶活性升高(高露姣等,2005;惠天朝等,2000)。本試驗中豬油組和混合油組的谷丙轉氨酶含量顯著高于其他各組(P<0.05),混合油組的谷草轉氨酶含量顯著高于其他各組 (P<0.05),這表明豬油組和混合油組魚體肝臟受到一定程度的損傷,豬油和魚油-豆油-豬油(3:4:3)不適宜作為大菱鲆的飼料脂肪源。這與高露姣等(2005)在飼料中添加豬油飼喂施氏鱘的研究結果一致。
4 結論
本試驗結果表明,魚油-豆油(1:1)、豆油和菜籽油可作為大菱鲆飼料合適的脂肪源,而豬油、混合油組魚體肝臟可能受到一定程度的損傷,豬油、混合油不適宜作為大菱鲆的飼料脂肪源。
