貴州禾來攏莖尖遺傳轉化體系的研究
2019-07-13 03:26:462
種子 2019年6期
2
(1.貴州大學生命科學學院/農業生物工程研究院/山地植物資源保護與種質創新教育部重點實驗室,貴州省農業生物工程重點實驗室, 貴陽 550025;2.貴州省農業科學院國家農業農村部DUS中心貴陽分中心, 貴陽 550006)
水稻(OryzasativaL.)不僅是重要的糧食作物,也是單子葉植物研究中最重要的模式作物之一。轉基因技術現已成為植物分子生物學研究基因功能的重要手段[1]。農桿菌介導法具有遺傳穩定性強、拷貝數較低且結合位點穩定等優勢,水稻轉基因植株絕大部分是利用農桿菌介導法獲得的[2]。以愈傷組織作為轉化受體需要對水稻苗幼胚或成熟種子在無菌條件下誘導獲得胚性愈傷組織后再進行轉化,這種方式需要嚴格的無菌環境,并且愈傷組織誘導及再分化對基因型依賴性較強,因此,不同水稻品種之間轉化難易程度差異較大[3,4,5]。以植物莖尖為受體進行遺傳轉化,無需經過愈傷組織誘導及再分化,具有轉化周期短、操作簡單、轉基因植物遺傳穩定等特點, 已在玉米、棉花、水稻等植物中成功應用[6-8]。前期對不同水稻品種的莖尖遺傳轉化體系轉化條件研究發現,不同水稻品種的最適條件各異,且主效因素也不一樣[6,9]。
來攏是從江地區特有的貴州香禾粳糯品種,前期品質測定發現其具有優良食味品質[10],可作為水稻品種改良的一種良好材料[11],但其對稻瘟病抗性較低,需對其進行品種改良。本實驗室曾經也以貴州禾來攏愈傷組織為受體,利用農桿菌侵染進行遺傳轉化并已獲得抗除草劑、抗稻瘟病的轉基因來攏植株,但轉化周期過長,且轉化效率較低[12,13]。因此,本研究以貴州禾來攏莖尖為材料設計正交試驗,研究不同菌液侵染濃度OD600、真空處理時間、真空滲透壓等因素對貴州禾來攏轉化效率及植株死亡率的影響,建立高效的農桿菌介導貴州禾來攏遺傳轉化體系,為基因工程改良貴州禾來攏奠定基礎。
1 材料與方法
1.1 材 料
植物材料為貴州禾來攏,農桿菌菌株為LBA 4404,均由貴州大學山地植物資源保護與種質創新教育部重點實驗室保存并提供。pCambia-Ubi-McCHIT1遺傳轉化載體由西南大學裴炎教授惠贈。
1.2 方 法
1.2.1遺傳轉化材料的準備
將貴州禾來攏成熟種子置于10 cm×12 cm網袋中清水浸泡,37 ℃避光培養,每8 h換水1次,待種子露白后,用清水清洗干凈。37 ℃避光催芽生根,待水稻胚芽鞘長至1.5~2.0 cm長時,切除莖環上部胚芽,暴露水稻的莖尖分生組織,用于農桿菌的侵染及轉化。
1.2.2農桿菌的活化及培養
取出-80 ℃保存的含有pCambia-Ubi-McCHIT1載體的農桿菌LBA 4404,接種到含有50 mg·L-1卡那霉素及20 mg·L-1利福平的YEP固體培養基上,28 ℃培養箱中暗培2 d。挑取單菌落于10 mLYEP液體培養基(卡那霉素及利福平濃度分別為50 mg·L-1、20 mg·L-1)中 180 r·min-1,28 ℃振蕩過夜培養。再取100μL菌液繼續放入新的YEP液體培養基中震蕩過夜,待菌液濃度OD600值達到0.8左右后,5 000 r·min-1離心收集菌體。
1.2.3遺傳轉化體系的建立
選擇農桿菌菌液侵染濃度OD600(0.4、0.6、0.8、1.0、1.2)、真空處理時間(2、5、8、11、14 min)及真空滲透壓(4、8、12、16、20 kPa)進行三因素五水平的正交試驗設計,設計為L25(53)正交表(詳見表1)。每個處理樣本為30粒,并做3次重復。AAM培養基[14]重懸農桿菌菌體并稀釋至相應濃度后,加入0.02%表面活性劑Silwet-L 77與100μmol·L-1乙酰丁香酮。將切除胚芽鞘的來攏種子浸泡在重懸后菌液中,使用Bio-Rad臺式基因槍對其進行真空處理,調節真空處理時間與真空處理壓強。處理完畢后,倒掉菌液,用吸水紙吸干后將水稻種子置于濕潤的珍珠巖上,黑暗條件下28 ℃共培養3 d。
表1 遺傳轉化正交設計與結果
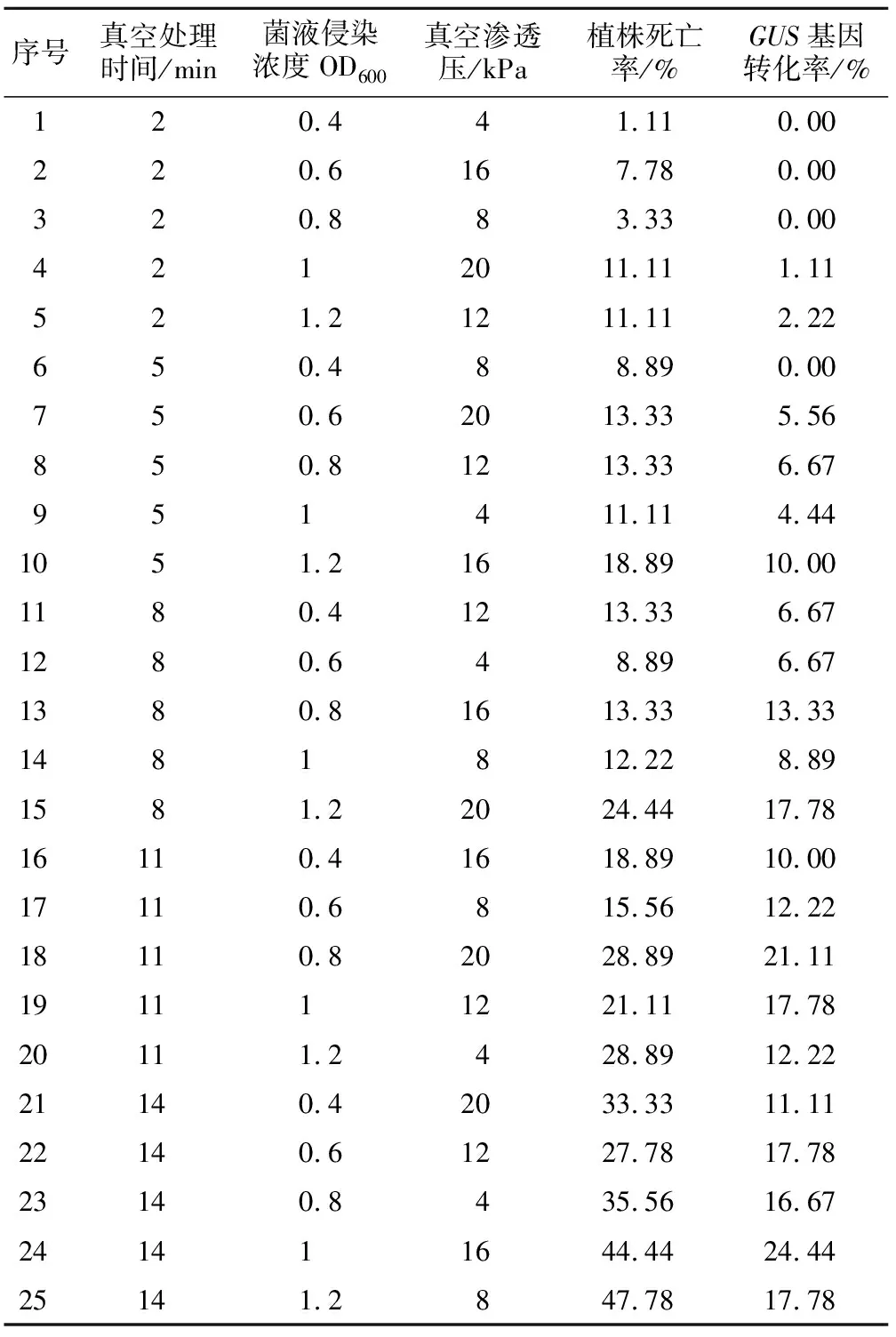
序號真空處理時間/min菌液侵染濃度OD600真空滲透壓/kPa植株死亡率/%GUS基因轉化率/%120.441.110.00220.6167.780.00320.883.330.004212011.111.11521.21211.112.22650.488.890.00750.62013.335.56850.81213.336.67951411.114.441051.21618.8910.001180.41213.336.671280.648.896.671380.81613.3313.331481812.228.891581.22024.4417.7816110.41618.8910.0017110.6815.5612.2218110.82028.8921.11191111221.1117.7820111.2428.8912.2221140.42033.3311.1122140.61227.7817.7823140.8435.5616.67241411644.4424.4425141.2847.7817.78
1.2.4植株死亡率及GUS組織化學檢測統計
共培3 d后,恢復正常光照,待水稻幼苗自然生長5 d,統計水稻幼苗死亡數;參照Jefferson[15]的方法,剪取新萌發的幼苗葉片進行GUS組織化學檢測,統計GUS染色陽性株數。計算植株死亡率和GUS基因轉化率。
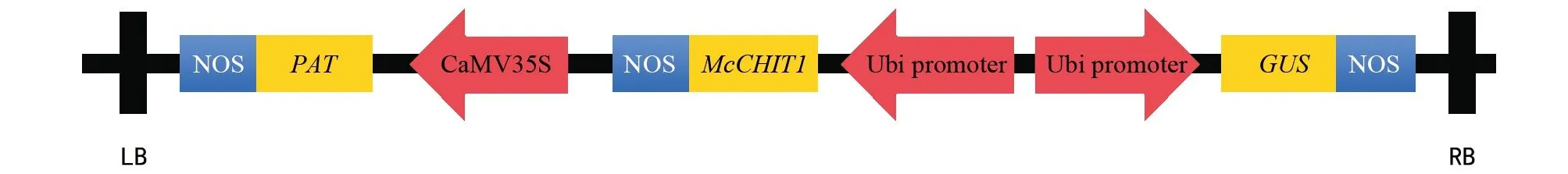
圖1 pCambia-Ubi-McCHIT1載體結構示意圖
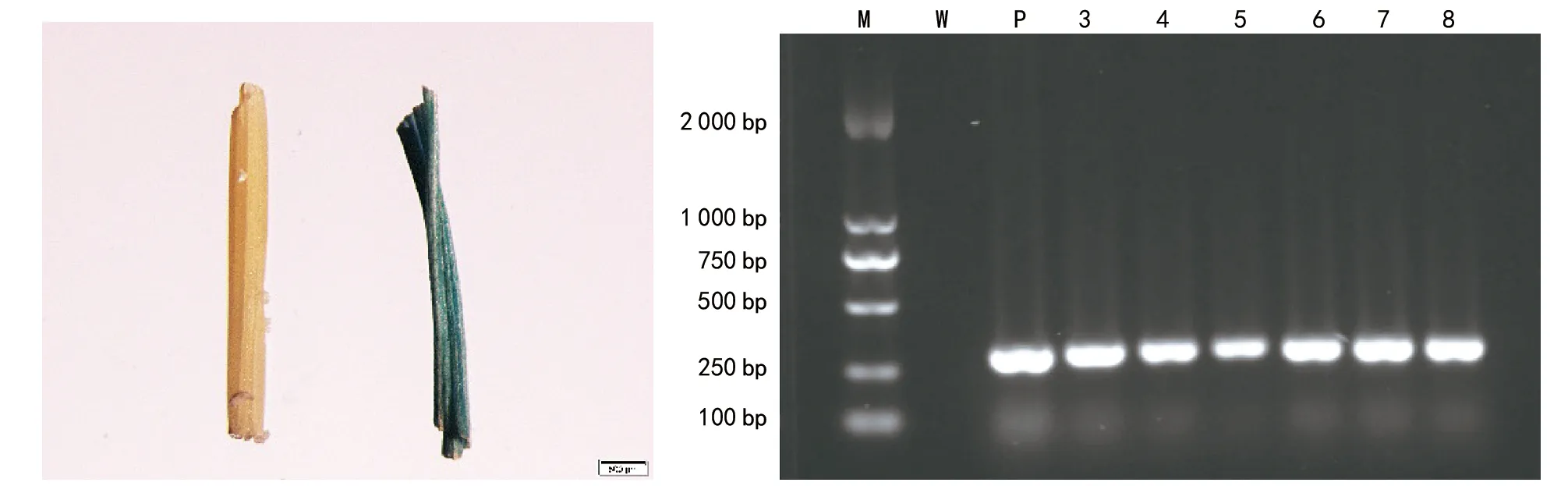
注:A為GUS組織染色(左為貴州禾來攏野生型;右為GUS陽性植株。bar=500 μL);B為轉基因植株PCR檢測(M為Marker;W為貴州禾來攏野生型;P為菌液DNA;3~8為轉基因植株)圖2 轉基因植株組織化學染色及PCR檢測
植株死亡率(%)=(植株死亡株數/總侵染樣本數)×100%;
GUS基因轉化率(%)=(GUS染色陽性植株/總侵染樣本數)×100%。
1.2.5轉基因植株的PCR檢測
將GUS染色陽性植株移栽至土壤中,常規肥水管理。按天根新型植物DNA提取試劑盒說明書提取來攏野生型及GUS染色陽性水稻葉片DNA,進行PCR檢測。PCR的反應條件為:94 ℃,3 min;94 ℃,30 s,54 ℃,30 s,72 ℃,30 s,35次循環;72 ℃延伸7 min;4 ℃保存。GUS特異性引物:Forward primer:5′-GGTGATTGATGAAACTGCTG-3′;Reverse primer:5′-GAACATTACATTGACGCAGG-3′,產物長度為304 bp。
1.2.6數據處理
試驗數據用SPSS 21.0軟件及Microsoft office Excel 2016軟件處理分析,多重比較選用鄧肯氏法。
2 結果與分析
2.1 各因素對貴州禾來攏死亡率的影響
通過統計貴州禾來攏死亡株數,計算出正交試驗中各組植株的死亡率,分析各因素對水稻死亡率的影響。結果表明,真空處理時間、菌液侵染濃度OD600、真空滲透壓3個因素越大,植株死亡率最高(表1)。方差分析結果發現,真空處理時間與菌液侵染濃度對來攏的死亡率均有顯著影響,真空滲透壓對死亡率影響不大(表2)。極差分析發現,真空處理時間的極值(R值)最大達到30.89,菌液侵染濃度次之,最小的是真空滲透壓。說明真空處理時間對植株死亡率的影響最大,真空滲透壓影響最小(表3)。
表2 3個因素對植株死亡率的方差分析
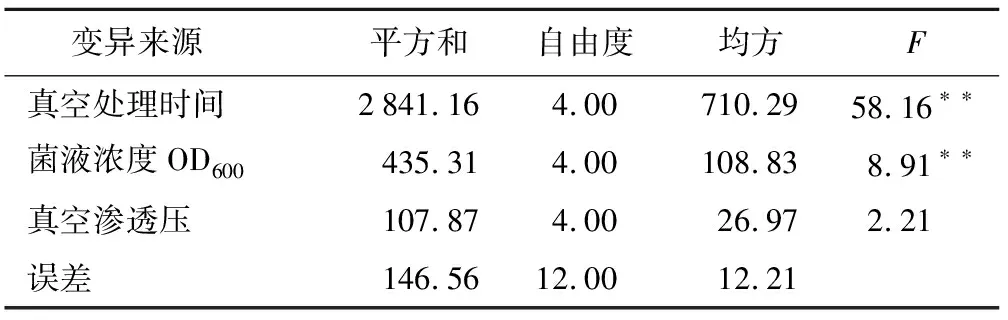
變異來源 平方和自由度均方F真空處理時間2841.164.00710.2958.16??菌液濃度OD600435.314.00108.838.91??真空滲透壓107.874.0026.972.21誤差146.5612.0012.21
表3 3個因素對植株死亡率的極差分析
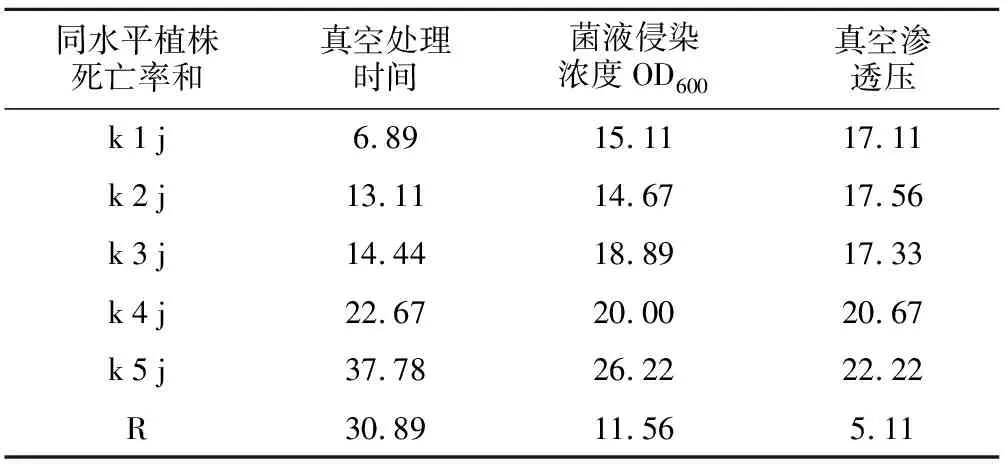
同水平植株死亡率和真空處理時間菌液侵染濃度OD600真空滲透壓k1j6.8915.1117.11k2j13.1114.6717.56k3j14.4418.8917.33k4j22.6720.0020.67k5j37.7826.2222.22R30.8911.565.11
2.2 各因素對貴州禾來攏遺傳轉化率的影響
用于遺傳轉化的pCambia-Ubi-McCHIT1載體具有內含子GUS報告基因,轉化成功的水稻植株經過GUS組織化學染色會呈現出藍色(如圖2 A)。對GUS組織化學染色陽性植株與來攏野生型的葉片進行PCR擴增,以質粒DNA為陽性對照,來攏野生型為陰性對照。結果表明:GUS染色陽性植株與質粒DNA均能得到大小304 bp左右的條帶,野生型來攏沒有條帶,說明外源基因已經成功轉入了GUS染色陽性植株(如圖2 B)。
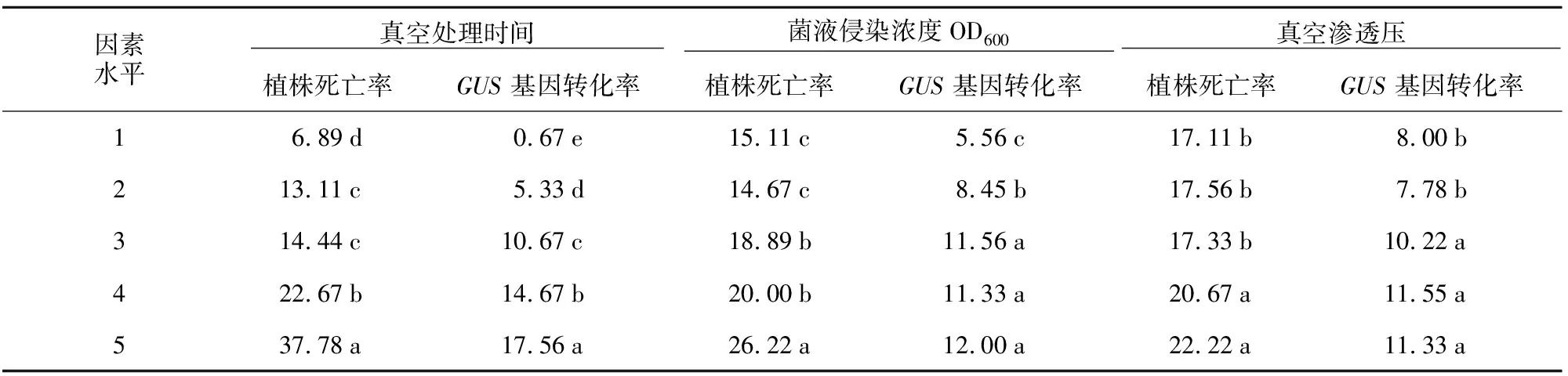
表6 各因素不同水平對植株死亡率及GUS基因轉化率的影響
因素水平 真空處理時間 菌液侵染濃度OD600 真空滲透壓 植株死亡率GUS 基因轉化率植株死亡率GUS 基因轉化率植株死亡率GUS 基因轉化率16.89d0.67e15.11c5.56c17.11b8.00b213.11c5.33d14.67c8.45b17.56b7.78b314.44c10.67c18.89b11.56a17.33b10.22a422.67b14.67b20.00b11.33a20.67a11.55a537.78a17.56a26.22a12.00a22.22a11.33a
注:表中同列不同小寫字母表示p<0.01差異顯著。
對GUS染色檢測呈陽性的植株數目統計分析,計算得出GUS基因轉化率,并對其進行極差分析與方差分析,結果表明:真空處理時間14 min,菌液侵染濃度OD6001.2,真空滲透壓16 kPa植株GUS轉化率最高(表1);方差分析表明,3個因素均對植株遺傳轉化率有顯著影響(表4);3個因素的R值大小順序為真空處理時間>菌液侵染濃度OD600>真空滲透壓,說明對水稻轉化率影響最大的是真空處理時間,其次為菌液侵染濃度,真空滲透壓對轉化率的影響最小(表5)。
表4 3個因素對GUS基因轉化率的方差分析
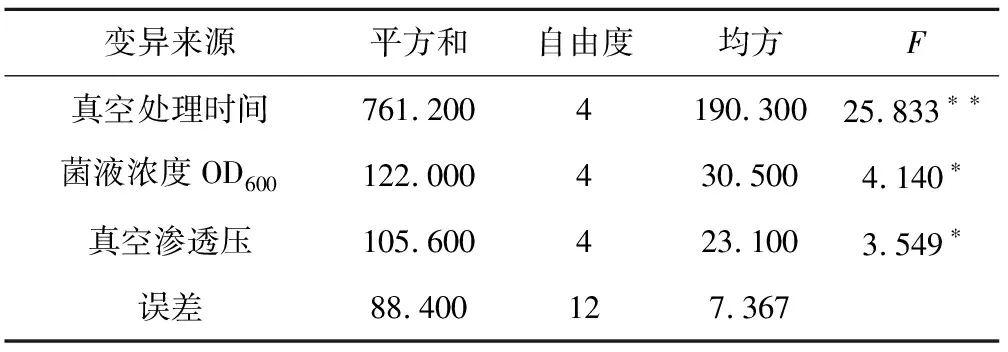
變異來源平方和自由度均方F真空處理時間761.2004190.30025.833??菌液濃度OD600122.000430.5004.140?真空滲透壓105.600423.1003.549?誤差88.400127.367
注:“*”表示p<0.05差異顯著,“**”表示p<0.01差異極顯著。
表5 3個因素對GUS基因轉化率的極差分析

同水平GUS基因轉化率和真空處理時間菌液侵染濃度OD600真空滲透壓k1j0.675.568.00k2j5.338.457.78k3j10.6711.5610.22k4j14.6711.3311.55k5j17.5612.0011.33R值16.896.443.78
2.3 各因素的不同水平對植株死亡率及轉化效率的綜合影響
隨著真空處理時間的增長,植株死亡率呈現出增加趨勢。處理5~8 min時,植株的死亡率并未出現顯著增加;處理時間大于8 min后,死亡率達到了20%以上,在14 min時達到最大值(為37.78%)。菌液侵染濃度OD600增大,植株死亡率也逐漸增加。當菌液濃度OD600達到1.2時,植株死亡率最大(為26.22%)。真空滲透壓對植株死亡率影響較小,真空滲透壓低于12 kPa時,植株死亡率無明顯變化(在17%左右);高于16 kPa后,死亡率明顯提高(達到20%以上)。GUS轉化率與真空處理時間成正相關。真空處理時間2 min時,GUS轉化率最小(僅達到0.67%);在高于8 min后,GUS轉化率能達到10%以上。菌液濃度OD600越大,GUS基因轉化率也越大,菌液濃度OD600為1.2時,GUS基因轉化率達到最高水平(為12%)。對真空滲透壓與GUS基因轉化率結果分析發現,滲透壓壓強低于8 kPa時,GUS基因轉化率均處于10%以下;大于12 kPa時,GUS轉化率達到10%以上,真空滲透壓16 kPa時基因轉化率最大(為11.55%)(表6)。
綜合各因素對植株死亡率及轉化效率的影響,認為:真空處理11 min,菌液侵染濃度OD6000.8,真空滲透壓16 kPa是農桿菌介導貴州禾來攏莖尖遺傳轉化的最優參數。
3 討 論
研究表明,在農桿菌介導的遺傳轉化中,外植體類型、菌液濃度、真空滲透壓以及真空處理時間等因素對植株的轉化效率及死亡率有密切的影響[16-18]。菌液濃度作為影響農桿菌轉化效率的關鍵因素之一,其最適濃度會隨著材料及方法的不同而發生很大的變化[19]。以植物愈傷組織為轉化受體的最適菌液濃度較低,在0.5左右;以莖尖為轉化受體的最適菌液濃度偏高,可以達到0.9以上[20,21]。張笑寒等認為,粳型黑糯米“平塘黑糯”與貴州禾“黎平雜邊禾“進行莖尖遺傳轉化體系構建中發現,菌液濃度與遺傳轉化率呈正相關[9];黃仁權等研究表明,菌液濃度對黎平雜邊禾死亡率影響較大但對平塘黑糯的影響不大,推測是由于水稻種類不同導致的[6]。本研究中貴州禾來攏與黎平雜邊禾的結果類似,菌液濃度對死亡率及轉化率均有顯著影響。本實驗室在前期研究發現:黑糯具有較白糯更強的抗逆性,可能是由于黑糯含有花青素等抗氧化性較強的物質,對生物和非生物脅迫的耐性都強。因此我們推測,黑糯米中由于含有花青素等物質,對農桿菌濃度的耐受性較高;白糯米對菌液濃度更敏感。
作為輔助農桿菌浸染植物的方法,真空滲透處理也會對植株的遺傳效率產生影響[22]。處理壓強過高、處理時間過長都會增大植株死亡率;處理壓強過低及處理時間過短,則會讓導入受體的農桿菌數量過少,降低基因的轉化率。真空處理時間是影響貴州禾來攏死亡率及轉化率的最關鍵因素,處理時間越長,來攏的死亡率與轉化率均越高。真空滲透壓壓強對來攏死亡率影響較小,以16 kPa為分界點,滲透壓壓強大于16 kPa后植株死亡率顯著提高到20%以上。來攏轉化率則以真空滲透壓12 kPa為分界點,高于12 kPa轉化率明顯升高。說明貴州禾來攏對真空處理具有一定適應力,短時間的高滲透壓對來攏的影響并不大,但長時間的處理會對來攏造成十分嚴重的影響。
