脲酶抑制劑與硝化抑制劑對稻田土壤硝化、反硝化功能菌的影響
2019-07-13 03:34:40張文學王少先夏文建劉增兵李祖章劉光榮
植物營養(yǎng)與肥料學報 2019年6期
張文學,王少先,夏文建,孫 剛,劉增兵,李祖章,劉光榮
(江西省農業(yè)科學院土壤肥料與資源環(huán)境研究所/農業(yè)部雙季稻營養(yǎng)與農業(yè)環(huán)境觀測實驗站/國家紅壤改良工程技術研究中心,南昌 330200)
尿素是一種高濃度的酰胺態(tài)氮肥,因其較高的含氮量被作為農業(yè)生產(chǎn)上最常用的氮肥,但是尿素施入農田后會導致氮素損失是一直被關注的。硝化-反硝化作用是稻田氮素損失的重要途徑[1],而且硝化、反硝化一般是緊密相連、相繼發(fā)生的,又被稱為成對硝化和反硝化過程[2],這兩個反應過程被證明均由土壤微生物所驅動[3]。
硝化作用可分為兩步:氨氧化過程 (NH3→NO2-)與亞硝酸鹽氧化過程 (NO2-→NO3-),氨氧化過程被認為是硝化作用的限速步驟[3-7]。氨氧化過程又可以分兩步為NH3→NH2OH→NO2-,且分別由氨單加氧酶(ammonia monooxygenase,AMO) 與羥胺氧化還原酶(hydroxylamine oxidoreductase,HAO) 催化完成[8]。氨單加氧酶是一種胞內酶,由基因amoA、amoB和amoC編碼的三個亞基組成,由于amoA基因的序列具有一定的保守性,且所編碼的蛋白亞基含有該酶的活性位點,因此常被作為氨單加氧酶的功能標記基因[4-5]。氨氧化細菌 (ammonia oxidizing bacteria,AOB)、氨氧化古菌 (ammonia-oxidizing archaea,AOA) 作為氨氧化的主要完成者,均含有編碼氨單加氧酶的基因amoA、amoB和amoC[9],且amoA基因被作為研究AOB與AOA的標記基因。反硝化作用(NO3-→NO2-→NO→N2O→N2) 一般發(fā)生在嫌氣或低氧的環(huán)境,硝酸鹽在硝酸鹽還原酶 (Nar)、亞硝酸鹽還原酶 (Nir)、NO還原酶 (Nor) 和N2O還原酶 (Nos) 的催化作用下依次還原為NO2-、NO、N2O、N2,對編碼以上四種酶的功能基因主要有narG、nirK/nirS、norB、nosZ[3,10],由于亞硝酸鹽還原酶 (Nir) 催化的反應是反硝化過程中關鍵的限速步驟,因此,關于編碼該酶的功能基因nirK和nirS的研究也較多,主要用作反硝化微生物群落的分子標記物[11],而且,研究發(fā)現(xiàn)nirK基因對環(huán)境因子的響應較nirS基因更靈敏[12-13]。
由于脲酶抑制劑可以通過抑制脲酶活性以達到延緩尿素水解為銨態(tài)氮的目的,硝化抑制劑可以抑制銨態(tài)氮的硝化作用,從而減少NO3-的形成、淋失以及隨后的反硝化損失[14-15],二者常被用在農業(yè)生產(chǎn)上提高氮肥利用率[16-17]。有報道指出,參與硝化、反硝化菌的群落結構以及豐度對土壤養(yǎng)分、pH、溫度等理化性狀的變化有著靈敏的響應[3,18-19]。但關于兩種抑制劑及其配施對稻田硝化與反硝化菌群結構、豐度的影響報道較少。本文以南方紅壤稻田為研究對象,在添加脲酶抑制劑NBPT與硝化抑制劑DMPP之后,研究了分蘗期與孕穗期土壤中銨態(tài)氮、硝態(tài)氮含量、amoA與nirK基因豐度以及AOB、AOA與反硝化細菌群落結構的變化,旨在進一步揭示抑制劑的作用機理及其對土壤環(huán)境的影響。
1 材料與方法
1.1 試驗地點
試驗于2012年4—7月在江西省農業(yè)科學院土壤肥料與資源環(huán)境研究所南昌試驗基地 (28°21′12″N,115°54′25″E) 進行。該區(qū)屬于亞熱帶濕潤氣候,海拔高度20 m,平均氣溫17.8℃,≥ 10℃的積溫為5432℃,無霜期長達240~307 d,年降水量1662 mm,降水季節(jié)分配不均,全年降水50%以上集中在4—6月。供試土壤為第四紀紅粘土發(fā)育的潮砂泥田,土壤質地為壤土。播種前土壤有機質含量20.12 g/kg,全氮含量2.06 g/kg、硝態(tài)氮含量1.8 mg/kg、銨態(tài)氮含量16.2 mg/kg、有效磷2.6 mg/kg、速效鉀94.55 mg/kg、pH 4.97、土壤容重為1.19 g/cm3。
1.2 供試材料與試驗設計
水稻供試品種為株兩優(yōu)30,脲酶抑制劑為NBPT (N-丁基硫代磷酰三胺),硝化抑制劑為DMPP(3,4-二甲基吡唑磷酸鹽)。供試肥料品種:氮肥為尿素 (含N 46%)、磷肥為鈣鎂磷肥 (含P2O512%)、鉀肥為氯化鉀 (含K2O 60%)。
試驗共設5個處理:1) 不施氮肥 (CK);2) 尿素(U);3) 尿素 + 脲酶抑制劑 (U + UI);4) 尿素 + 硝化抑制劑 (U + NI);5) 尿素 + 脲酶抑制劑 + 硝化抑制劑 (U + UI + NI)。設3次重復,各小區(qū)隨機區(qū)組排列,每個小區(qū)面積30 m2(5 m × 6 m),各小區(qū)以50 cm的分隔行隔開,且保持小區(qū)間的田埂高出地面40 cm,并用塑料薄膜包裹田埂以防止相互串水串肥,實現(xiàn)各小區(qū)獨立排灌的管理目的。試驗氮 (N)、磷(P2O5)、鉀 (K2O) 用量分別為 135 kg/hm2、75 kg/hm2和150 kg/hm2。抑制劑與尿素混勻施入,抑制劑用量為尿素的1%。氮肥和磷肥作基肥于移栽前一次性施入;鉀肥分3次施入,40%作基肥,30%作分蘗肥,30%作孕穗肥。試驗于2012年3月10日播種,4月23日移栽,7月20日收獲,水稻種植密度以及各項栽培管理措施同當?shù)剞r民習慣。
樣品采集:在水稻分蘗期、孕穗期分別采集各小區(qū)耕層土壤,一部分保存于4℃冰箱用于測定土壤銨態(tài)氮、硝態(tài)氮含量,另一部分保存于-20℃用于土壤DNA提取,分析AOB、AOA以及反硝化細菌的基因豐度與群落結構。
1.3 樣品測定與數(shù)據(jù)分析
1.3.1 土壤中銨態(tài)氮與硝態(tài)氮含量的測定 土壤中的銨態(tài)氮與硝態(tài)氮含量用1 mol/L的KCl溶液浸提,采用全自動間斷化學分析儀 (Smartchem TM200 discrete chemistry analyzer) Smartchem 200測定。
1.3.2 土壤微生物總DNA的提取 將土壤樣品按照土壤DNA提取試劑盒 (Fast DNA SPIN Kit for soil MP-bio,USA) 的說明進行操作,并用0.7% (w/v) 的瓊脂糖檢驗DNA,提取成功的樣品保存于-20℃冰箱待用。
1.3.3 聚合酶鏈式反應 (PCR) 對提取成功的DNA模板進行聚合酶鏈式反應 (polymerase chain reaction,PCR),運用特異性引物對土壤中特定微生物的DNA片段進行擴增。AOB與AOA的amoA基因所用的擴增引物分別為amoA-1F/amoA-2R[20]與Crenamo A23f/Crenamo A616r[21-22],反硝化細菌的nirK基因所用的引物為F1aCu/R3Cu[23],具體引物序列以及PCR反應程序見表1。PCR反應體系共25 μL:12.5 μL 的 2 × EasyTaq PCR SuperMix (TransGen 生物技術公司,中國北京),0.5 μL的上、下引物和1 μL稀釋10倍的DNA模板,10.5 μL的滅菌雙蒸水,終體積為25 μL。在MyCycler熱循環(huán)儀 (Bio-Rad公司)進行PCR反應,所有PCR產(chǎn)物采用1%的瓊脂糖凝膠電泳檢驗PCR擴增反應的效果。
1.3.4 熒光定量PCR AOB、AOA的amoA基因和nirK基因的定量PCR利用熒光定量PCR儀Bio-Rad iQ5 (Bio-Rad,USA) 分析,運行程序按定量試劑盒說明書執(zhí)行。PCR體系包括:5 μL SYBR Green I PCR Mix (TaKaRa,Dalian,China),0.2 μL 的引物,0.8 μL DNA 模板,終體積為10 μL。AOB、AOA的amoA基因和nirK基因定量PCR所用引物見表1。含有目標基因片段的質粒從108—102進行10倍濃度梯度連續(xù)稀釋制作標準曲線,通過溶解曲線確定定量PCR的擴增特異性;每個樣品重復3次。反應結束后,運行程序會根據(jù)標準曲線以及樣品的Ct值(每個樣品管內的熒光信號到達設定閾值時所經(jīng)歷的循環(huán)數(shù)) 定量計算出樣品的起始拷貝數(shù)。

表1 PCR引物序列及反應程序Table 1 Primers and conditions used for the conventional PCR
1.3.5 變性梯度凝膠電泳 (DGGE) 聚丙烯酰胺濃度為8% (w/v),AOB、AOA與nirK型反硝化細菌的DGGE變性濃度分別為50%~70%、30%~50%與45%~65%。在1倍TAE緩沖液中,75 V電壓、60℃條件下電泳16 h,電泳后用1 μL的SyBR green I (Invitrogen) (稀釋10000倍) 核酸染料染色30 min,然后用Bio-Rad成像系統(tǒng) (Bio-Rad,USA)拍照。DGGE圖譜采用Quantity One軟件分析樣品的電泳條帶特征,聚類分析采用UPGMA運算法則。
1.3.6 序列比較與系統(tǒng)發(fā)育分析 將DGGE圖譜中的優(yōu)勢菌群切膠回收純化,并用表1中相應的引物進行PCR擴增,將擴增產(chǎn)物進行純化后測序[生工生物工程 (上海) 股份有限公司],測序得到的序列信息輸入NCBI (www.ncbi.nlm.nih.gov) 通過Blast與Gene Bank中的已知序列比對,確定序列的微生物種類,并運用MEGA6.0軟件的鄰接法 (Neighbor joining method) 進行系統(tǒng)發(fā)育分析。
1.3.7 數(shù)據(jù)處理 數(shù)據(jù)經(jīng)Excel 2007整理后,采用SAS9.1軟件進行統(tǒng)計分析;PCR-DGGE成像用quantity one軟件分析。
2 結果與分析
2.1 土壤銨態(tài)氮與硝態(tài)氮含量
稻田土壤中的銨態(tài)氮含量明顯高于硝態(tài)氮的含量(圖1)。在水稻分蘗期,與處理U相比,處理U +UI和U + UI + NI的銨態(tài)氮含量顯著降低,而處理U +NI與處理U相比無顯著差異,這可能由于NBPT減緩了尿素水解,顯著降低了此時土壤的銨態(tài)氮含量,而單獨添加DMPP則無此效應。在孕穗期,所有處理的土壤銨態(tài)氮含量較分蘗期的均急劇下降,可能是由于分蘗期到孕穗期水稻快速生長對氮肥的大量吸收所致;施氮肥的四個處理相比,添加NBPT的處理U + UI與U + UI + NI的銨態(tài)氮含量明顯高于處理U與U + NI的,這可能由于NBPT此時已分解,失去了對脲酶活性的抑制作用,尿素進一步水解,增加了銨態(tài)氮的含量,可為水稻的后期生長提供充足的氮源。
與土壤銨態(tài)氮含量相比,硝態(tài)氮含量極低,且同一時期內處理間的差異均不顯著 (P> 0.05),這說明添加NBPT雖然顯著降低了分蘗期土壤中銨態(tài)氮的含量,但是對于硝態(tài)氮含量沒有顯著影響,這可能由于淹水條件下硝化作用極弱,導致銨態(tài)氮的含量對硝化作用無明顯影響,同理,硝化抑制劑DMPP對硝化反應的影響也很微弱。
2.2 土壤硝化、反硝化菌豐度
處理間AOB的amoA基因拷貝數(shù)存在顯著差異(圖2)。在分蘗期,AOB的amoA基因拷貝數(shù)范圍在2.0 × 106~3.4 × 106copies/g,施氮處理中 AOB 的amoA基因拷貝數(shù)顯著高于不施氮處理;處理U +NI與U + UI + NI的amoA基因拷貝數(shù)顯著低于處理U和U + UI。這說明施用氮肥顯著增加了分蘗期稻田土壤中AOB的豐度,然而添加硝化抑制劑DMPP能夠有效抑制AOB的生長。在孕穗期,所有施氮處理相比CK均增加了土壤amoA基因拷貝數(shù),而施用氮肥4個處理間無顯著差異,說明施用氮肥對孕穗期的AOB生長依然有促進作用,但DMPP對AOB的抑制作用已消失,可能此時DMPP的時效性已過。
圖3顯示,AOA的amoA拷貝數(shù)范圍在17.3 ×106~20.5 × 106copies/g之間,是氨氧化細菌amoA基因拷貝數(shù)的6.1~10.2倍,說明在紅壤稻田AOA在數(shù)量上居于主導地位;AOA的amoA基因拷貝數(shù)在分蘗期明顯高于孕穗期的,說明生育期對AOA的豐度有明顯影響,而所有處理間始終無顯著差異,說明AOA的豐度對施用氮肥、NBPT以及DMPP均無明顯響應。
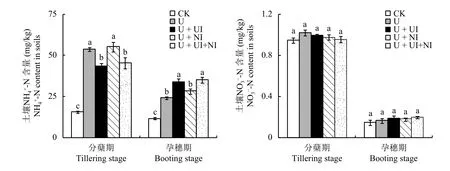
圖1 土壤銨態(tài)氮與硝態(tài)氮含量Fig.1 Contents of NH4+-N and NO3--N in soils
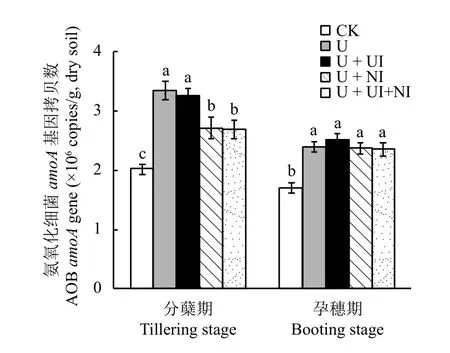
圖2 土壤氨氧化細菌 (AOB)amoA基因拷貝數(shù)Fig.2 Ammonia-oxidizing bacterial (AOB)amoAgene copy numbers in soils
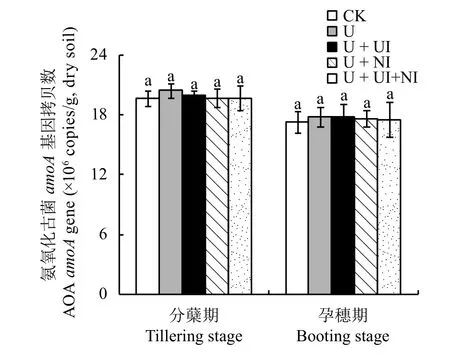
圖3 土壤氨氧化古菌 (AOA)amoA基因拷貝數(shù)Fig.3 Ammonia-oxidizing archaeal(AOA)amoAgene copy numbers in soils

圖4 土壤反硝化細菌nirK基因拷貝數(shù)Fig.4 Denitri fi ernirKgene copy numbers in soils
由圖4可知,nirK基因拷貝數(shù)在21.0 × 107~36.3 × 107copies/g之間,在分蘗期與孕穗期,施氮肥處理的nirK基因拷貝數(shù)均顯著高于不施氮肥的,而處理U、U + UI、U + NI與U + UI + NI在分蘗期或孕穗期均無顯著差異,說明施用氮肥可以顯著提高nirK型反硝化細菌的豐度,而脲酶抑制劑NBPT或硝化抑制劑DMPP對此無明顯影響。分蘗期的nirK基因拷貝數(shù)均顯著高于孕穗期的,可能主要原因是孕穗期土壤中氮肥的消耗導致反硝化細菌的響應,另外,不同生育期土壤生態(tài)環(huán)境的變化對此也存在有一定程度的影響。從三種菌群豐度來看,古菌amoA/細菌amoA拷貝數(shù)比值大于6.1,而nirK/(細菌amoA+ 古菌amoA) 拷貝數(shù)比值大于11.1,這可能由于稻田特定的生態(tài)環(huán)境所決定。
2.3 土壤銨態(tài)氮、硝態(tài)氮含量與硝化、反硝化菌豐度的相關性
由表2可知,AOB的豐度與分蘗期銨態(tài)氮、硝態(tài)氮以及孕穗期的銨態(tài)氮含量呈極顯著正相關 (P<0.01),說明AOB對氮肥的施入有靈敏的響應,而且對氨氧化作用有重要的貢獻;另外,因為NBPT對土壤NH4+-N含量有明顯的調節(jié)作用,因此推測NBPT對AOB的豐度也有著間接的影響。AOB的豐度與孕穗期的硝態(tài)氮含量相關性不顯著,可能由于此時硝態(tài)氮含量過低所致。
AOA豐度與兩個時期的土壤銨態(tài)氮、硝態(tài)氮含量的相關性均不顯著 (P> 0.5),表明AOA比較穩(wěn)定,對氮肥的施入沒有明顯響應。
反硝化細菌與分蘗期銨態(tài)氮、硝態(tài)氮以及孕穗期的銨態(tài)氮含量呈極顯著正相關 (P< 0.01),這與AOB的規(guī)律相似。以上結果說明,AOB與反硝化細菌對氮肥的施入反應靈敏,而AOA則比較穩(wěn)定。
2.4 硝化、反硝化菌的群落結構特征
運用變性梯度凝膠電泳 (DGGE) 的方法研究AOB的群落結構。首先利用引物amoA-1F/amoA-2R對AOB的amoA基因進行PCR擴增,擴增產(chǎn)物再通過DGGE分析,得到相應的DGGE圖譜 (圖5)。與不施氮肥相比,施用氮肥對分蘗期與孕穗期AOB的群落結構均有明顯影響,主要表現(xiàn)為優(yōu)勢條帶明顯增多。在分蘗期,CK處理的主要條帶包括B3和B4,施用氮肥的4個處理還明顯增強了B1和B2條帶的信號強度;然而,單施氮肥處理與添加了抑制劑的處理 (U與U + NI、U + UI、U + UI + NI)之間AOB的圖譜差異不大,且條帶B2和B3信號強度均較高,表明B2與B3是施氮肥稻田土壤中AOB的優(yōu)勢菌群。此外,我們發(fā)現(xiàn)不同生育期對AOB群落結構的影響在處理CK的差異比較明顯,分蘗期處理CK的優(yōu)勢條帶為B3,而在孕穗期優(yōu)勢條帶轉變?yōu)锽2,表明隨著時間的推移,土壤AOB優(yōu)勢菌群是動態(tài)變化的,這可能與氣候條件變化或者土壤養(yǎng)分的消耗有關。優(yōu)勢菌群B3在孕穗期時,所有處理的信號均明顯變弱,這可能由于條帶B3所代表的AOB對氮肥的響應更靈敏,隨氮肥的減少,B3豐度明顯下降。經(jīng)聚類分析 (UPGMA) 的結果表明,所有處理分為兩大類,分蘗期的CK處理和孕穗期的CK單獨一簇,其余處理聚為一簇,兩個時期所有施用氮肥處理的AOB群落結構的相似度大于68%,表明施用氮肥顯著影響了AOB的群落結構,而脲酶抑制劑NBPT和硝化抑制劑DMPP對AOB群落結構的影響較小。
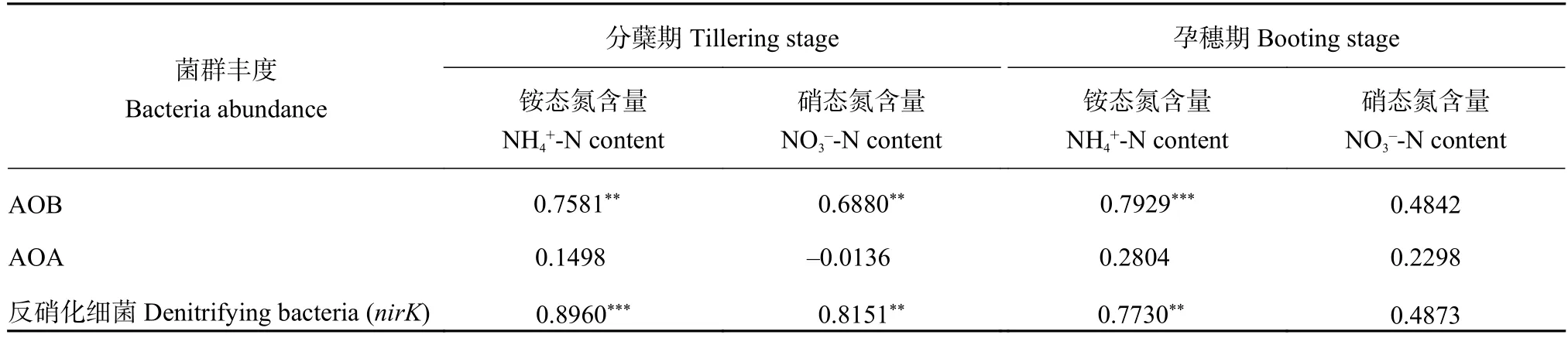
表2 土壤銨態(tài)氮、硝態(tài)氮含量與硝化、反硝化菌豐度的相關性Table 2 Relations between contents of NH4+-N and NO3--N with abundances of AOB, AOA and denitrifying bacteria in soils
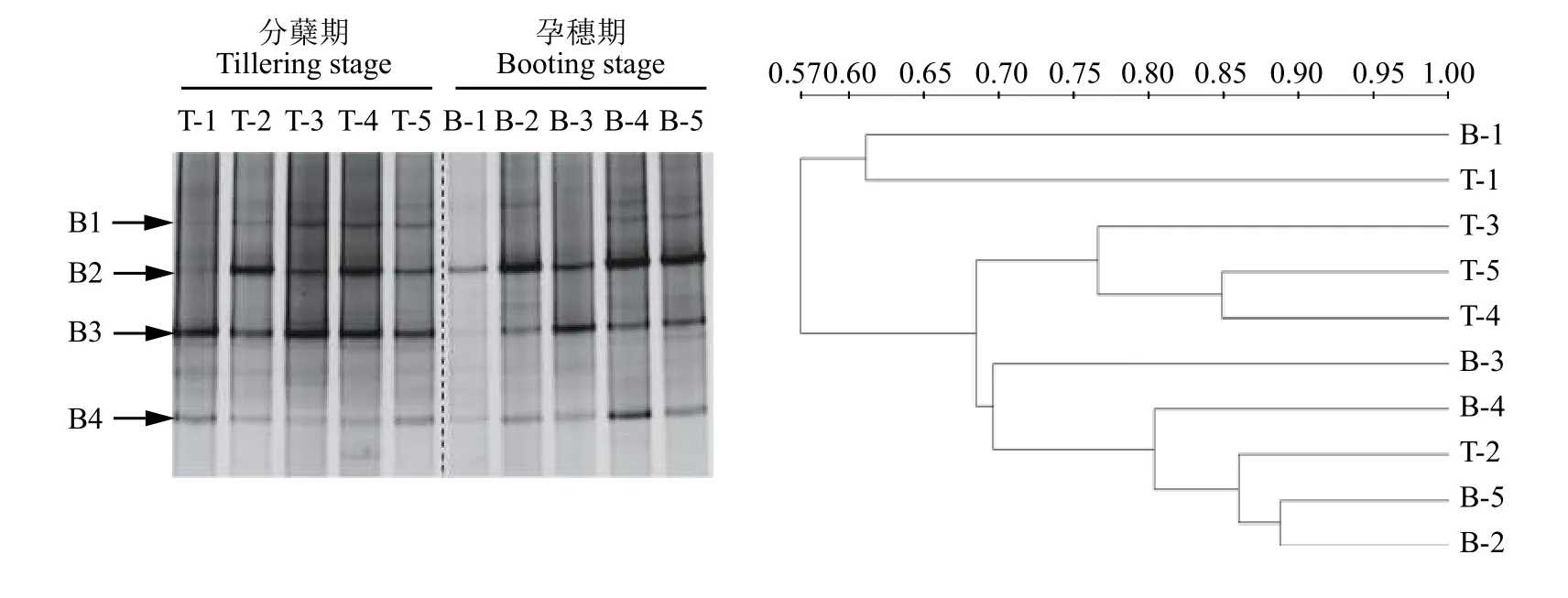
圖5 氨氧化細菌amoA基因的DGGE圖譜及其聚類分析Fig.5 DGGE profile and clustering analysis of DGGE patterns of ammonia-oxidizing bacterialamoAgene
對AOB的DGGE凝膠上的4個條帶進行切膠純化測序,所得序列經(jīng)NCBI數(shù)據(jù)庫的BLAST比對分析后發(fā)現(xiàn),所有序列與數(shù)據(jù)庫中來源于土壤的氨氧化細菌amoA基因的相似性大于96%。將檢測到的4個基因序列及其相似序列構建系統(tǒng)發(fā)育樹 (圖6)可知,B1與B3聚為一簇,二者同源性較高,B2與B4各自為一簇;與B1和B3同源性較高的序列有來源于砂壤土的亞硝化單胞菌目 (Nitrosomonadales)以及來源于紅壤、酸性土壤、湖泊的氨氧化細菌amoA基因;與B2同源性較高的序列有來源于土壤的亞硝化螺菌屬 (Nitrosospirasp.) 以及來源于稻田氨氧化細菌的amoA基因;與B4同源性較高的序列有來源于土壤的亞硝化螺菌屬以及亞硝化葉菌屬(Nitrosolobus multiformis)。
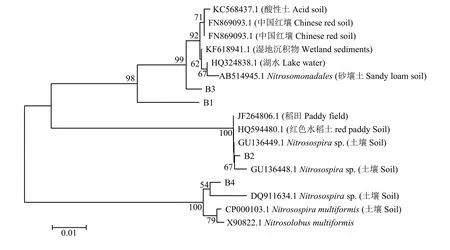
圖6 amoA基因的系統(tǒng)進化樹Fig.6 Phylogenetic tree of ammonia-oxidizing bacterialamoAsequences from DGGE bands using neighbor-joining analysis
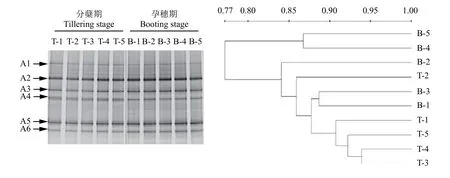
圖7 氨氧化古菌amoA基因的DGGE圖譜及其聚類分析Fig.7 DGGE profile and clustering analysis of ammonia-oxidizing archaealamoAgene
利用引物crenamoA 23fd和crenamoA 616rd對AOA的amoA基因進行PCR擴增,隨后進行DGGE分析,相應的DGGE圖譜見圖7。所有處理均包括6條主要的條帶 (A1—A6),說明這6條帶是稻田土壤中AOA的優(yōu)勢菌群。相比AOB的群落結構變化,各處理對AOA的群落沒有明顯影響;兩個時期間也無顯著變化,說明該稻田土壤中AOA對不同施肥處理以及環(huán)境變化不敏感,在水稻不同生育期間較為穩(wěn)定。聚類分析 (UPGMA) 的結果表明,兩個時期所有處理的氨氧化古菌群落結構的相似度大于77%,說明施用氮肥、脲酶抑制劑NBPT以及硝化抑制劑DMPP對AOA的群落結構均無明顯影響。
對AOA的DGGE凝膠上的6個條帶進行切膠純化測序,所得序列經(jīng)NCBI數(shù)據(jù)庫的BLAST比對分析后發(fā)現(xiàn),與數(shù)據(jù)庫中AOA的amoA基因相似度高于92%,將目的序列及其相似的序列建立系統(tǒng)發(fā)育樹 (圖8),所有條帶被分為兩大類:條帶A2、A5與A3、A6聚為一簇,A1、A4聚為一簇,說明A2、A5、A3、A6序列同源性較高,A1與A4的序列同源性較高;與A2、A5同源性較高的序列主要來自于酸性土壤,與A3、A6同源性較高的序列主要來源于紅壤以及稻田AOA的amoA基因;而與A1、A4聚為一簇的主要來自于玉米地及沉積物AOA的amoA基因等。
利用引物F1aCu/R3Cu對反硝化細菌的nirK基因進行PCR擴增,并進行DGGE分析 (圖9),所有處理在分蘗期的條帶數(shù)明顯多于孕穗期的,其中條帶K1、K2、K3、K5與K7信號強度最為強烈,說明包含這5類nirK基因的細菌是該稻田土壤中nirK型反硝化菌的優(yōu)勢菌群;施用氮肥以及不同抑制劑對分蘗期的反硝化細菌群落結構沒有明顯影響。在孕穗期,所有處理的各條帶均明顯變弱,尤其是不施肥處理下,可見條帶更為稀疏,說明此時nirK型反硝化細菌的豐度明顯下降,這可能與此時氮肥已大量消耗有關,因為反硝化細菌對氮肥有靈敏的響應[12]。聚類分析 (UPGMA) 的結果表明,兩個時期所有處理的反硝化細菌群落結構被分為三大類:孕穗期的CK處理單獨一簇,其余處理的聚為一簇,分蘗期的所有處理聚為一簇,說明不同生育期對nirK型反硝化菌群落結構有著明顯的影響。以上結果說明,水稻生育期以及施用氮肥對nirK型反硝化菌群落結構有明顯影響,而兩種抑制劑則無此效應,這說明兩種抑制劑的作用具有專一性,對反硝化細菌無明顯影響。
對反硝化細菌nirK基因DGGE凝膠上的7個條帶進行切膠測序分析,所得序列經(jīng)NCBI數(shù)據(jù)庫的BLAST比對分析,建立系統(tǒng)發(fā)育樹 (圖10),系統(tǒng)發(fā)育樹明顯的分為兩簇,大多數(shù)條帶 (K1、K2、K3、K4、K5和K6) 分布在第一簇中,這一簇的nirK基因序列主要來源于農田、高山草甸、稻田等,而K7單獨分布在另一簇,與其它nirK基因遺傳距離較遠。
3 討論
脲酶抑制劑可以提高氮素利用率[16,24],本研究表明,添加NBPT (或NBPT與DMPP配施) 延緩了尿素水解,顯著提高了水稻孕穗期土壤中的銨態(tài)氮含量,為水稻后期生長提供充足氮肥,這可能是其增產(chǎn)增效的主要原因,而單獨添加DMPP則無此效應,說明稻田不適宜單獨添加硝化抑制劑。

圖8 氨氧化古菌amoA基因的系統(tǒng)進化樹Fig.8 Phylogenetic tree of ammonia-oxidizing archaealamoAsequences from DGGE bands using neighbor-joining analysis
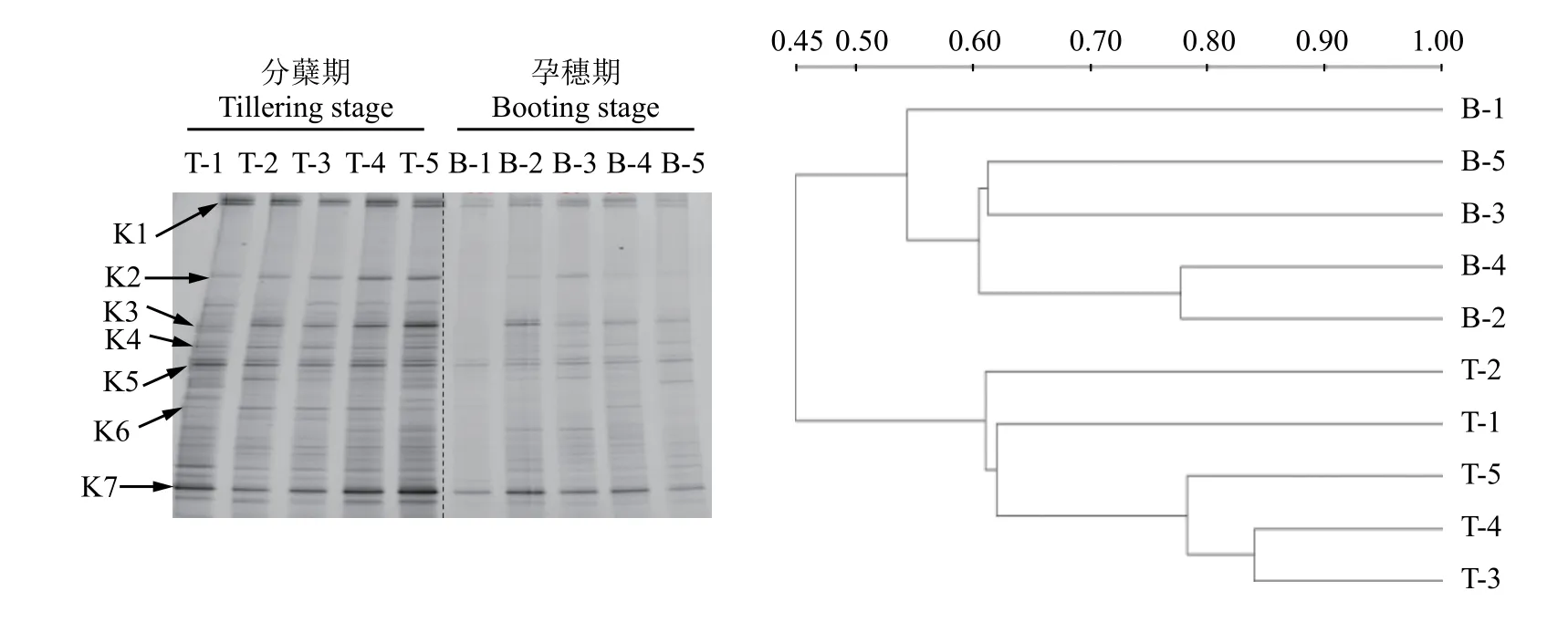
圖9 反硝化細菌nirK基因的DGGE圖譜及其聚類分析Fig.9 DGGE profile and clustering analysis of denitrifying bacterialnirKgene
有研究表明,土壤養(yǎng)分、pH以及施肥制度等環(huán)境因素對AOB、AOA的生長繁殖有不同程度的影響[25-28]。農田施入氮肥可明顯促進AOB的生長[29],增加其豐度[30-31],而對AOA則無明顯影響[32-33]。不同的硝化抑制劑對AOB以及AOA的影響不同,有的可顯著抑制AOB的生長[34-37],對AOA沒有明顯影響[38];有的則效果相反[39-40],這可能由于不同抑制劑的作用機理不盡相同所致,因為抑制劑可通過抑制硝化菌的生長、呼吸[16,41-42]或活性[16]等多種方式來發(fā)揮作用。本研究表明,施用氮肥顯著增加了分蘗期與孕穗期稻田土壤中AOB的豐度,然而添加硝化抑制劑DMPP在分蘗期能夠有效抑制AOB的生長,到孕穗期對AOB的抑制作用消失,可能此時DMPP已降解所致。DMPP對AOA的生長則始終無明顯的影響,這可能與本試驗地的稻田土壤性質以及生態(tài)環(huán)境有關,或者由于AOA有較強的穩(wěn)定性而導致。由此可知,DMPP在稻田減弱硝化反應的主要途徑是通過抑制AOB的生長而實現(xiàn)。添加NBPT對AOA、AOB的豐度無明顯影響,證明NBPT僅與土壤中的脲酶結合起作用,抑制其活性,對土壤微生物無明顯的直接影響。
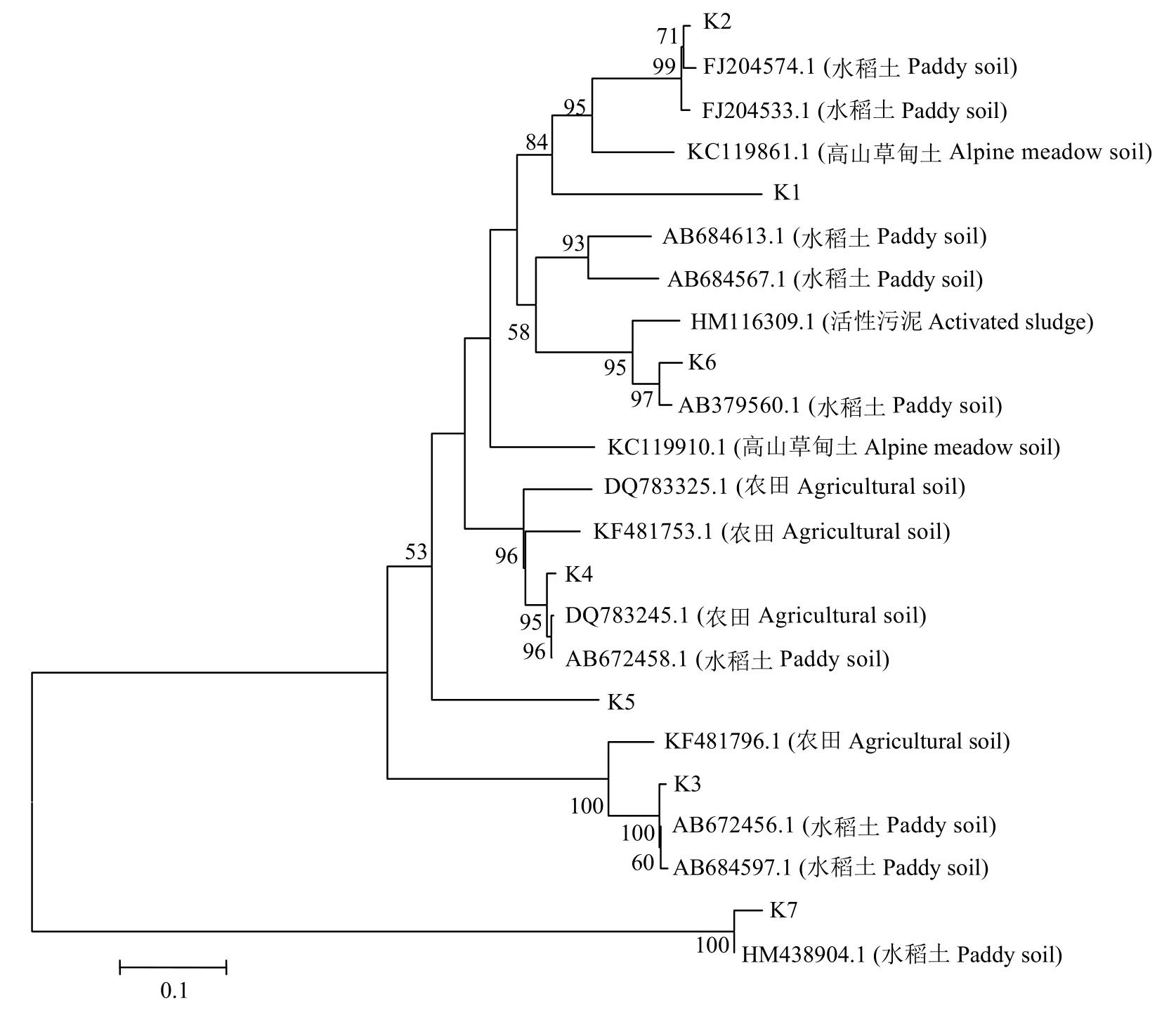
圖10 反硝化細菌nirK基因的系統(tǒng)進化樹Fig.10 Phylogenetic tree of denitrifying bacterialnirKgene based on DGGE band sequences using neighbor-joining analysis
土壤理化性質如土壤養(yǎng)分、含水量、pH、有機質等因素同樣影響著反硝化細菌的群落結構和種群豐度[13,43]。有研究表明,施用氮肥可以明顯提高反硝化細菌nirK、nirS基因的豐度[12,44-47]。本研究結果證明,施用氮肥可顯著提高nirK型反硝化細菌的豐度,隨著氮肥消耗,到孕穗期,所有處理下反硝化細菌的豐度明顯下降,而添加抑制劑NBPT與DMPP的則沒有表現(xiàn)出明顯的變化,這說明氮肥可直接引起反硝化細菌的響應,不同生育期則可能通過土壤理化性質或土壤微生態(tài)的變化間接地影響反硝化細菌的豐度,而抑制劑則無此效應,進一步說明NBPT與DMPP對土壤環(huán)境的安全性。有報道指出,DMPP可通過降低底物濃度而間接影響nirK型反硝化菌的豐度[48],本研究沒有監(jiān)測到此種關聯(lián),可能由于所試稻田長期淹水,反硝化反應底物濃度極低所致。
有報道指出,在水稻田等pH較低的土壤中,AOA的數(shù)量遠超過AOB的[49-50],且在氨氧化過程中發(fā)揮主導作用[38],同時,反硝化細菌nirK基因的拷貝數(shù)又遠超過amoA(AOA + AOB) 基因的拷貝數(shù)[11]。本研究的結果也表明,古菌amoA/細菌amoA拷貝數(shù)比值大于6.1,而nirK/(細菌amoA+ 古菌amoA) 拷貝數(shù)比值大于11.1,與以上結論一致。
從三種菌群豐度與銨態(tài)氮、硝態(tài)氮含量的相關性分析來看,AOB與nirK反硝化細菌的豐度與分蘗期時期土壤中銨態(tài)氮、硝態(tài)氮含量以及孕穗期銨態(tài)氮含量呈極顯著正相關,證明了二者對氮肥的敏感性,而AOA則沒有此相關性,說明AOA的穩(wěn)定性,與Chen等報道一致[32]。由于NBPT對銨態(tài)氮含量有明顯的調節(jié)作用,由此推測,NBPT對AOB以及反硝化菌也有間接的影響。
施用化肥對土壤硝化、反硝化菌群落結構及其組成有著明顯的影響[19]。本研究中DGGE指紋圖譜表明,施用氮肥明顯增加了圖譜中AOB的條帶數(shù),隨著氮肥的消耗,反應靈敏的優(yōu)勢菌群到了孕穗期信號強度明顯減弱;施用氮肥對反硝化細菌群落結構的影響,主要體現(xiàn)在氮肥大量消耗后的孕穗期,此時,不施氮肥處理的DGGE圖譜中優(yōu)勢菌群條帶僅剩3條依稀可辨,遠遠少于施氮肥處理的;AOA在兩個時期對氮肥均沒有明顯的響應。另外,抑制劑的施用在DGGE圖譜中也沒有表現(xiàn)出明顯影響,可能由于DGGE圖譜只是定性地反應群落組成,而抑制劑只對菌群豐度有影響,對其組成沒有影響所致。
4 結論
1) NBPT顯著提高了分蘗期土壤銨態(tài)氮含量,顯著降低了孕穗期的銨態(tài)氮含量,AOB與nirK反硝化細菌的豐度與土壤中銨態(tài)氮、硝態(tài)氮含量有極顯著正相關關系,AOA則與兩種形態(tài)的氮相關性不顯著,說明AOA比較穩(wěn)定,AOB與反硝化細菌對氮肥有靈敏的響應,尤其是無機氮離子,而NBPT可調節(jié)銨態(tài)氮含量,因此,NBPT對AOB與nirK反硝化細菌的豐度可能有著間接的影響。
2) 硝化抑制劑DMPP對AOB有明顯的抑制作用,但僅表現(xiàn)在分蘗期,孕穗期抑制作用基本消失,而對AOA以及反硝化細菌則始終沒有明顯影響,DMPP在稻田抑制硝化作用的途徑是通過抑制AOB的生長來實現(xiàn)的。
3) DGGE圖譜直觀地反映了三種菌群對氮肥的響應,AOB與反硝化細菌的群落結構在施用氮肥下條帶數(shù)明顯增加,而AOA則無明顯響應,再次證明AOB與反硝化細菌對氮肥的敏感性以及AOA的穩(wěn)定性。
4) 脲酶抑制劑NBPT對三類微生物的群落結構以及豐度沒有明顯的直接影響,硝化抑制劑DMPP僅在分蘗期對AOB的生長有抑制作用,這說明NBPT與DMPP對土壤生態(tài)的安全性。
