香豆素對入侵植物節節麥種子萌發及幼苗生長的影響
2019-07-24 08:19:30昌孝濤袁美麗
浙江農林大學學報 2019年4期
關鍵詞:質量
王 寧,昌孝濤,袁美麗,陳 浩
(1.河南科技大學 林學院,河南 洛陽 471023;2.洛陽市林業工作站,河南 洛陽 471000;3.洛陽市隋唐城遺址植物園,河南 洛陽471002)
化學除草劑的危害愈來愈受到人們重視,利用植物化感作用對田間雜草進行生態防控被認為是環境友好,促進農業可持續發展的新技術,并已被列入當代農業生態學研究的重要研究方向[1-2]。香豆素(C9H6O2)是香豆素類物質中形式最簡單的化合物。鄔彩霞等[3]采用色譜分離等技術,發現黃花草木樨Melilotus officinalis中化感物質含量最高的是香豆素。黃花草木樨對藜Chenopodium album[4],稗草Echinochloa crusgalli[5], 蘇丹草Sorghum sudanense[6]和多花黑麥草Lolium multiflorum[7]等多種雜草均存在較強的抑制作用。此外,覃逸明等[8]研究發現:在鳳丹Paeonia ostii根際土壤中存在的5種化感物質中,香豆素抑制作用最強。CHON等[9]研究發現:與肉桂酸、香草酸等其他酚酸類化感物質相比,香豆素對苜蓿Medicago sativa的化感抑制作用最強。RAZAVI等[10]研究發現:當香豆素質量濃度達100 mg·L-1時,可完全阻礙生菜Lactuca sativa種子萌發及其幼苗根、莖生長。上述研究均表明:香豆素有作為新型除草劑的開發利用潛力,而有關香豆素對入侵植物的化感作用研究尚未見報道。節節麥Aegilops tauschii為禾本科Gramineae山羊草屬Aegilops植物,最早起源于西亞及東歐等地,現已成為世界惡性雜草[11]。1955年,節節麥在中國河南省新鄉地區首次被發現,如今已廣泛入侵至渭河流域陜西、甘肅,黃河流域河北、河南、山東以及內蒙古等地,并成為中國麥田中最難防除的惡性雜草之一[12-13]。據報道,僅2003年河北南部地區由于節節麥危害導致的小麥損失就達10%~25%[14]。因嚴重威脅糧食的生產與安全,早在2007年節節麥已被列入中國首次發布的《中華人民共和國進境植物檢疫性有害生物名錄》[15]。生產上僅有甲基二磺隆和異丙隆對節節麥具有一定防除效果,但由于不同小麥品種及節節麥種群對上述2種除草劑的敏感性存在差異[16-17],導致它們在實際生產中的推廣應用受到一定限制。這也是節節麥入侵范圍愈來愈廣泛的重要原因之一。為此,本研究以入侵植物節節麥為試材,探討香豆素對其種子萌發及幼苗生長的影響,以期為節節麥的生物防控提供參考。
1 材料與方法
1.1 材料
2017年5月,節節麥種子來源于河南省新鄉市農業科學院試驗田(35°18′N,113°52′E)。研究在河南科技大學開元校區林學院園林植物實驗室進行。固體香豆素來源于上海試劑廠。
1.2 方法
香豆素溶液配置:準確稱取(萬分之一天平)75 mg香豆素,蒸餾水定容至500 mL,充分溶解后得150 mg·L-1香豆素溶液,分別稀釋得到50,100 mg·L-1的溶液,置4℃冰箱保存。使用時提前取出,室內放置3~5 h,避免低溫溶液對種子萌發造成影響。
種子萌發試驗:采用培養皿沙培法[18]。選取籽粒飽滿、大小基本一致的節節麥種子,先用體積分數為1%的次氯酸鈉溶液浸泡10 min,之后蒸餾水反復沖洗,室內自然晾干備用。將種子置于鋪有經高溫消毒處理石英砂的培養皿(直徑12 cm)內,加入10 mL(經預試驗確定)處理液,50粒·皿-1,利用透明保鮮膜覆蓋包裹培養皿(保證培養皿口保鮮膜平展)以減少試驗過程中水分的散失,并稱量記錄每皿的質量(試驗期間根據石英砂顏色,加入適量處理液保持每皿質量不變)。各處理5次重復,各重復3皿。以加等量蒸餾水的處理為對照(ck)。置于晝夜25℃/15℃,光暗各12 h的光照培養箱內培養。每天觀察統計1次種子萌發情況(以胚根長度大于等于種子長度為萌發標準),培養10 d后各處理隨機選取10株幼苗分別測量根長及苗高。其中,發芽率RG=(10 d內供試種子發芽數/供試種子數)×100%,發芽指數IG=∑(Gt/Dt), 其中:Dt為相應發芽天數,Gt為逐日發芽種子數。 抑制率[4,19]RI=(T0-T1)/T0。 其中:T1為處理值,T0為對照值。RI<0為促進作用,RI>0,為抑制作用,RI絕對值大小代表抑制的強弱。
幼苗生長試驗:首先對節節麥種子進行催芽處理(水浸泡24 h,25℃培養箱內),選出露白基本一致的種子播種于裝有適量石英砂的塑料杯(開口直徑10 cm,高15 cm)內,10粒·杯-1,隨后加入15 mL(經預備實驗確定)不同質量濃度的處理液,以加等量蒸餾水的為對照(ck),各處理5次重復,各重復3杯。置于晝夜25℃/15℃,光暗各12 h的光照培養箱內培養。并分別在處理的5,10和15 d采樣,取全株進行各項生理指標的測定。電導法測定相對電導率(REC)[20];酸性茚三酮法測定脯氨酸質量分數,氮藍四唑光還原法測超氧化物歧化酶活性,硫代巴比妥酸法測定丙二醛質量摩爾濃度[21]。
1.3 統計方法
用Excel對數據進行初步處理,利用SPSS 18.0進行單因素方差分析,數據為平均值±標準差。
2 結果與分析
2.1 香豆素對節節麥種子萌發及幼苗生長指標的影響
隨香豆素溶液質量濃度的增加,節節麥種子發芽率、發芽指數均明顯下降(表1)。不同質量濃度處理的發芽率、發芽指數與對照差異均達到顯著水平(P<0.05)。150 mg·L-1處理中,發芽率僅有31.60%,較對照下降66.38%。此外,從抑制率可看出,發芽率及發芽指數的抑制率均大于0,且呈質量濃度效應的變化。由此表明,香豆素溶液對節節麥種子萌發存在較強的抑制作用。
隨香豆素質量濃度的增加,節節麥幼苗的苗高及根長同樣呈明顯的減少趨勢。其中,100和150 mg·L-1的處理苗高均顯著低于對照(P<0.05)。此外,不同質量濃度處理的根長均顯著低于對照(P<0.05)。香豆素對苗高及根長的抑制率均大于0,且也隨溶液質量濃度的增加而增大。由此表明,香豆素溶液對節節麥幼苗的生長也存在明顯抑制作用。

表1 香豆素溶液對節節麥種子萌發和幼苗生長過程中生長指標的影響Table 1 Coumarin solution influence of Aegilops tauschii growth indexes on seed germination and growth of seedlings
2.2 香豆素對節節麥幼苗生理指標的影響
由圖1A可知:相同質量濃度處理中,節節麥幼苗的相對電導率隨時間的延長呈快速上升的變化。其中,50 mg·L-1處理中,處理后第5天時,相對電導率略低于對照,之后則均高于對照,至第15天時較第5天時增加顯著(P<0.05)。100和150 mg·L-1處理中,相對電導率均高于對照,至第10天時較第5天時增加均已達到顯著水平(P<0.05)。
相同質量濃度處理中,丙二醛質量摩爾濃度隨時間的延長呈逐漸上升的變化(圖1B)。其中,50 mg·L-1處理中,至第15天時增加達到顯著水平(P<0.05)。100和150 mg·L-1處理中,至第10天時丙二醛質量摩爾濃度較第5天時增加已達到顯著水平(P<0.05)。同樣,相同時間下,丙二醛質量摩爾濃度與香豆素質量濃度呈正相關。
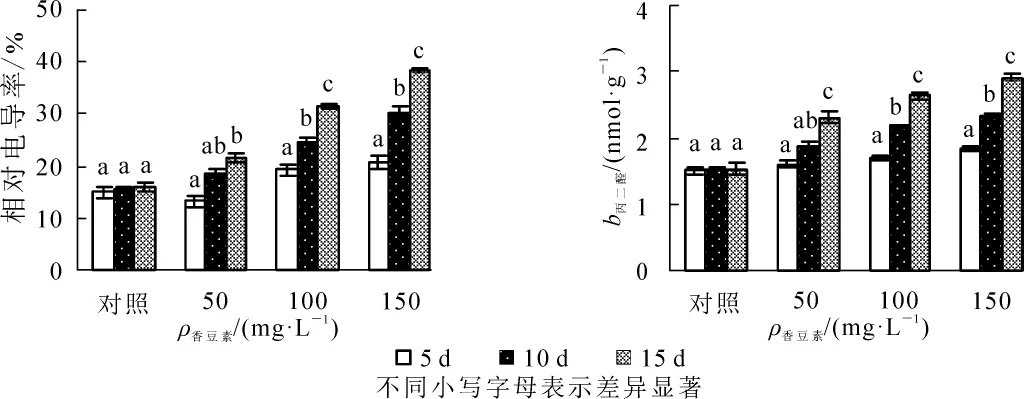
圖1 不同質量濃度香豆素對節節麥幼苗相對電導率及丙二醛質量摩爾濃度的影響Figure 1 Relative electric conductivity and MDA content of Aegilops tauschii seedling under different concentrations of coumarin solution
相同質量濃度處理中,節節麥幼苗的脯氨酸質量分數隨時間延長呈逐漸增加的變化(圖2A)。其中,不同質量濃度香豆素溶液處理中,至處理后第10天時脯氨酸質量分數較第5天時增加均已達到顯著水平(P<0.05)。150 mg·L-1處理中,至第15天時較第10天時增加未達到顯著水平。且相同時間下,脯氨酸質量分數與香豆素質量濃度呈正相關。
50和100 mg·L-1處理中,超氧化物歧化酶活性隨時間的延長呈持續增加的變化,而150 mg·L-1處理中,超過氧化物歧化酶活性呈先升后降的變化(圖2B)。其中,50及100 mg·L-1處理中,至處理后第10天時超氧化物歧化酶活性較第5天時增加已達到顯著水平(P<0.05)。150 mg·L-1處理中,至第10天時超氧化物歧化酶活性較第5天時增加顯著,至第15天時較第10天時略有下降,但差異不顯著。且相同時間下,超氧化物歧化酶活性與香豆素質量濃度呈正相關。
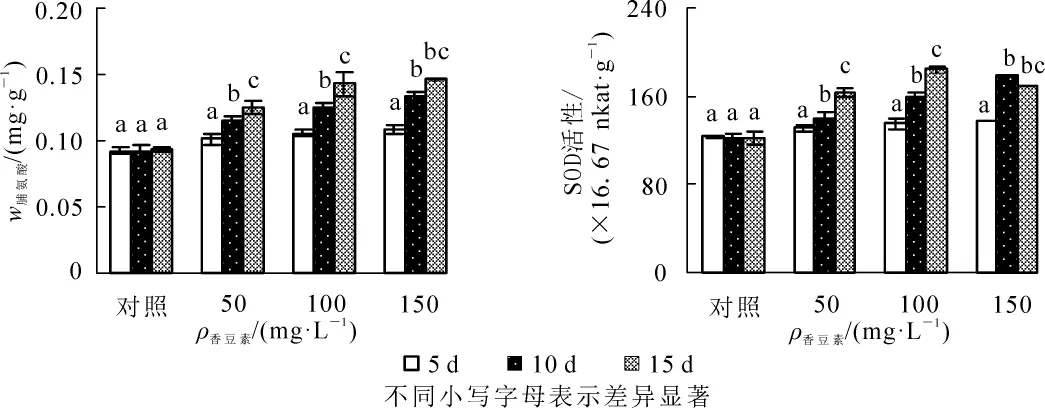
圖2 不同質量濃度香豆素對節節麥幼苗脯氨酸質量分數及超氧化物歧化酶活性的影響Figure 2 Proline content and SOD activity of Aegilops tauschii seedling under different concentrations of coumarin solution
3 討論
種子能否正常萌發決定了植物的繁衍及生存,對該物種的更新至關重要。而發芽率的增加及下降則會降低植物在群落中的競爭力及多度[22]。幼苗階段在植物的生命周期中較為關鍵,影響著植物的形態建成,且是植物對外界環境較為敏感的時期[23]。本試驗結果表明:香豆素溶液對節節麥種子萌發產生明顯的抑制作用,且抑制作用隨香豆素溶液質量濃度的增加而增大。在王婧怡等[6]對蘇丹草,姚丹丹等[7]對多花黑麥草的研究中均發現:香豆素溶液對上述2種植物種子萌發存在一定的化感抑制作用。香豆素溶液處理造成節節麥種子發芽率下降的原因可能有:一是香豆素可能影響了種子萌發所需關鍵性酶等物質,造成萌發過程中萌發所需能量物質的缺乏,從而導致種子活力的下降;二是香豆素可能改變了節節麥種皮透性,進而影響種子的吸水作用,造成種子內部物質的外滲,而此類物質造成了真菌病原體的生長,導致種子腐爛而無法萌發。本研究的種子萌發后期,也確有種子腐爛現象的發生。此外,香豆素溶液處理對節節麥幼苗的苗高及根長生長也存在質量濃度效應的化感抑制作用。其中,不同質量濃度處理中根長的抑制率均明顯大于苗高。由此也表明:香豆素對節節麥幼苗根長生長的化感抑制作用大于苗高,這可能與根系最先直接接觸并吸收到處理物質有關,只有在根系被抑制達到一定程度后,幼苗的地上部分才會逐漸受到影響。這與宋亮等[24]利用酚酸類物質對苜蓿Medicago sativa的研究結論相符。
相對電導率和丙二醛是植物質膜受傷害程度的重要反映指標。本研究中,相對電導率及丙二醛質量摩爾濃度均隨處理時間的延長呈逐漸增加的變化。其中,相對電導率的增加表明香豆素對節節麥幼苗膜結構造成一定破壞,導致電解質外滲的增大;而丙二醛質量摩爾濃度的增加則表明膜脂過氧化作用的逐漸增大。此外,50 mg·L-1溶液處理中,至第10天時相對電導率及丙二醛質量摩爾濃度較第5天時增加均不顯著。這可能是香豆素質量濃度低且處理時間短,節節麥通過自身的調節機制,對香豆素造成膜系統損傷進行了積極修復的結果。
滲透調節物質含量的增加是植物對逆境脅迫的重要生理反映,而游離脯氨酸是植物體內重要的滲透調節物質之一。本研究中,隨時間的延長,不同溶液處理中的脯氨酸質量分數均顯著增加,這是節節麥通過脯氨酸質量分數增加以適應香豆素脅迫的結果,這也與香豆素處理蘇丹草[6]、多花黑麥草[7]可溶性糖含量增加的結論相符。而150 mg·L-1處理中,至第15天時較第10天時增加不顯著,這可能與香豆素溶液的過度脅迫所致。超氧化物歧化酶是植物體內重要的保護酶,其活性大小是植物抗逆性強弱的重要體現。本研究中,隨時間的延長,超氧化物歧化酶活性呈持續增加的變化,表明節節麥啟動抗氧化系統以應對香豆素處理造成的其體內活性氧自由基的逐漸增加,這也是節節麥為適應香豆素脅迫而產生的應激反應。150 mg·L-1處理中,至第15天時較第5天時增加顯著,但較第10天時略有下降。這表明隨著香豆素脅迫的進一步加劇,幼苗體內活性氧自由基的增加速率逐漸超過了超氧化物歧化酶的清除能力,造成自由基不斷積累,最終導致超氧化物歧化酶活性的下降,這也與SCANDALIOS[25]及YE等[26]研究結論一致。
猜你喜歡
中學生數理化·中考版(2022年10期)2022-11-10 09:37:42
中學生數理化·八年級物理人教版(2022年12期)2022-02-14 07:08:42
中學生數理化·八年級物理人教版(2021年12期)2021-12-31 03:23:08
中學生數理化·中考版(2020年10期)2020-11-27 01:59:48
中國生殖健康(2019年2期)2019-08-23 08:12:08
石油化工建設(2018年6期)2018-04-22 03:16:54
產品可靠性報告(2017年7期)2017-09-05 09:49:12
中學生數理化·八年級物理人教版(2017年12期)2017-04-18 12:59:38
汽車觀察(2016年3期)2016-02-28 13:16:26
民生周刊(2014年7期)2014-03-28 01:30:54
