植物多功能調控因子SnRK2研究進展
2019-07-24 11:24:28苗麗麗劉秀林張宏紀毛新國景蕊蓮
麥類作物學報 2019年7期
關鍵詞:植物
苗麗麗,劉秀林,張宏紀,毛新國,景蕊蓮
(1.黑龍江省農業科學院博士后科研工作站,黑龍江哈爾濱 150086; 2.黑龍江省農業科學院作物資源研究所,黑龍江哈爾濱150086; 3.黑龍江省農業科學院大豆研究所,黑龍江哈爾濱 150086; 4.中國農業科學院作物科學研究所,北京 100081)
干旱、高鹽、極端溫度等非生物逆境極大地限制了植物生長,嚴重影響農作物生產。為了適應不利環境,植物在長期進化過程中形成了復雜的應對保護機制,如調控抗逆相關基因表達,以維持細胞結構和功能的完整性。蛋白激酶和蛋白磷酸酶共同參與的蛋白質可逆磷酸化是植物在逆境條件下調節體內能量代謝和信號轉導的重要方式。目前植物逆境脅迫應答過程中研究最多、涉及生理代謝與基因調控最廣的蛋白激酶家族主要有鈣依賴蛋白激酶CDPK、MAPK蛋白激酶和蔗糖非發酵相關蛋白激酶SnRK(sucrose non-fermenting 1-related protein kinase)[1]。SnRK屬于Ser/Thr蛋白激酶,是植物生長發育和脅迫應答調控網絡的樞紐[2]。根據其結構和功能不同,分為SnRK1、SnRK2、SnRK3三個亞家族。SnRK1主要參與碳代謝,如糖和淀粉的生物合成、碳水化合物在器官中的分配[3-4]。植物SnRK1與酵母SNF1(sucrose non-fermenting 1)和哺乳動物AMPK(腺苷酸活化蛋白激酶,AMP-activated protein kinase)在結構和功能上高度相似,被認為是真核生物的能量傳感器。SnRK2和SnRK3是植物特有的蛋白激酶亞家族,主要參與對滲透脅迫和逆境信號分子ABA(abscisic acid)的應答。
目前SnRK2的功能及其作用機理尚未完全解析。為了促進植物SnRK2基因家族的研究和利用,結合筆者所在課題組多年來對小麥SnRK2基因家族的研究結果,本文對SnRK2結構和分類、調控機制和生物學功能進行了綜述。
1 SnRK2的結構和分類
SnRK2是一個相對較小的蛋白激酶亞家族。在擬南芥、水稻、玉米、小麥和大豆等多種植物中均發現了SnRK2基因家族成員,不同物種中SnRK2成員數量不同(表1)。
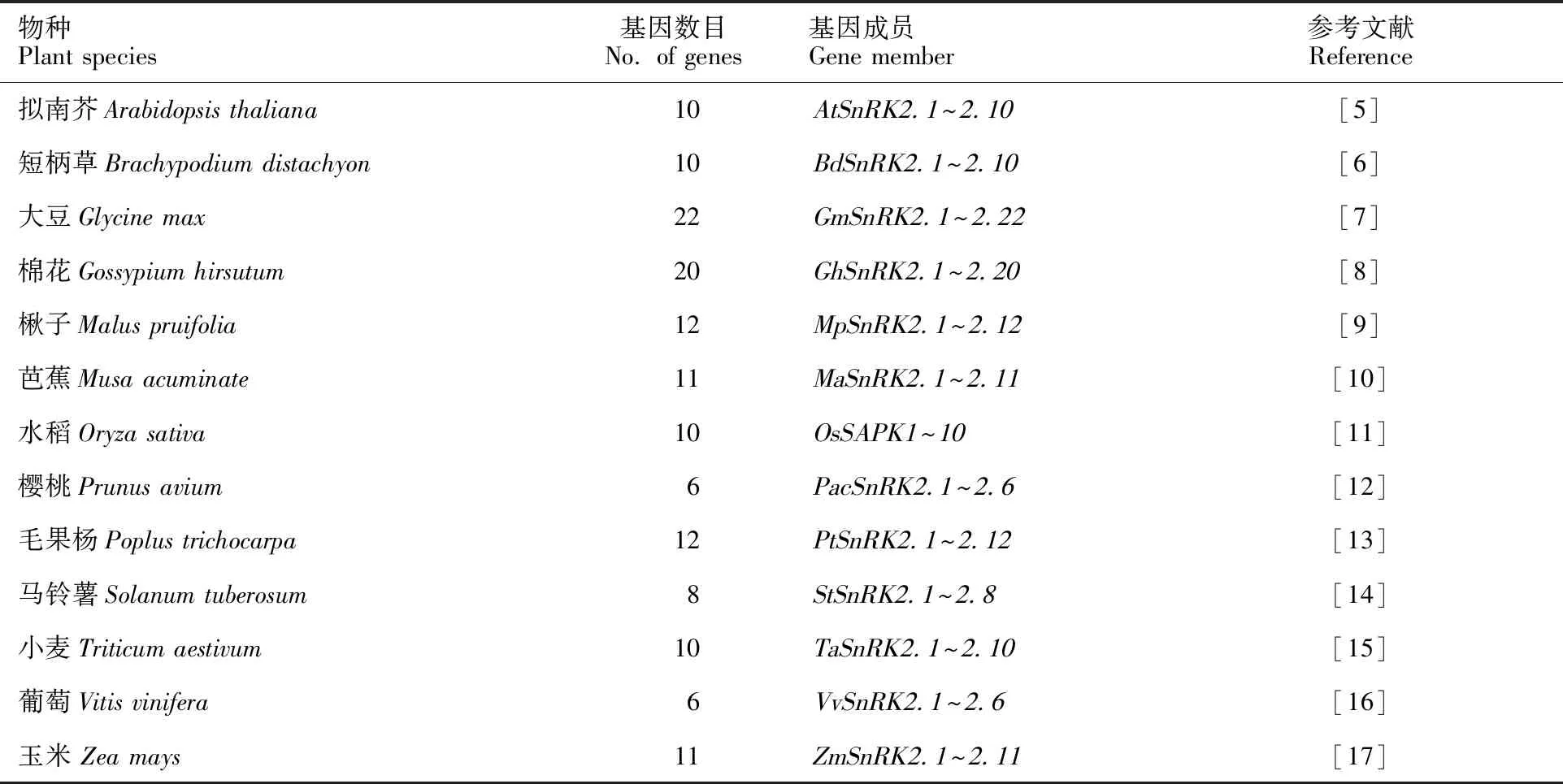
表1 不同植物中 SnRK2基因數目Table 1 The members of SnRK2 genes in different plant species
SnRK2蛋白分子量約為40 kD,含有兩個結構域: N端激酶結構域和C端調節結構域[18]。N端結構域異常保守,與AMPK和SNF1的激酶結構域高度同源,相似性達42%~46%; C端結構域又分為2個亞結構域Domain I 和Domain II。 Domain I由靠近N端激酶結構域后的30個氨基酸組成,存在于所有SnRK2成員中,主要受逆境脅迫激活,但不依賴ABA。Domain II由靠近C端的40個氨基酸組成,為ABA依賴型SnRK2所特有,是響應ABA應答的必須結構(圖1)。研究發現,SnRK2的C端結構域與酶激活性、ABA信號轉導及蛋白間互作有關[11]。

圖1 SnRK2的結構域[19]
目前有兩種分類方法,一種是根據C端酸性氨基酸不同,將SnRK2分為SnRK2a和SnRK2b,其中SnRK2a富含天冬氨酸,SnRK2b富含谷氨酸;另一種分類方法較為常用,依據是否受ABA誘導,將SnRK2分為3個群:GroupⅠ、Group Ⅱ和Group Ⅲ,其中GroupⅠ不受ABA誘導,Group Ⅱ受ABA微弱誘導,而Group Ⅲ受ABA強烈誘導。擬南芥10個SnRK2基因成員中,AtSnRK2.1、AtSnRK2.4、AtSnRK2.5、AtSnRK2.9和AtSnRK2.10屬于GroupⅠ;AtSnRK2.7和AtSnRK2.8屬于Group Ⅱ;AtSnRK2.2、AtSnRK2.3和AtSnRK2.6屬于Group Ⅲ[20]。而小麥的10個SnRK2基因成員與擬南芥無同源對應關系,TaSnRK2.4、TaSnRK2.5、TaSnRK2.6和TaSnRK2.7屬于GroupⅠ;TaSnRK2.1、TaSnRK2.2和TaSnRK2.3屬于Group Ⅱ;TaSnRK2.8、TaSnRK2.9和TaSnRK2.10屬于Group Ⅲ[15]。
2 SnRK2參與調控基因表達的機制
磷酸化對于激活SnRK2有促進作用,SnRK2主要通過磷酸化修飾來調控下游基因表達和蛋白質活性。目前有多個SnRK2磷酸化位點已被鑒定,最具代表性的是AtSnRK2.6的Ser175,其磷酸化狀態是激酶活性的關鍵,該磷酸化位點在蛋白激酶中普遍存在,如AtSnRK2.10和煙草SnRK2成員NtOSAK的Ser158是相似的磷酸化位點[21]。
SnRK2活性調控以自身磷酸化為基礎。目前對ABA依賴型SnRK2的可逆磷酸化已有比較深入的研究。ABA受體PYR/PYL/RCAR的發現是ABA信號通路研究的標志性進展[22]。在ABA依賴信號通路中,SnRK2蛋白激酶Group Ⅲ 成員是信號傳遞的樞紐,組成了ABA-PYR-PP2C-SnRK2-轉錄因子相偶聯的信號通路。缺乏ABA時,A類磷酸酶PP2C與SnRK2結合,此時SnRK2沒有活性;ABA存在時,ABA首先與其受體PYR/PYL/RCAR家族成員結合并引起受體構象變化,異構化的受體作用于PP2C的催化位點進而抑制其催化活性。ABA受體與PP2C結合促使后者釋放出SnRK2,后者通過自身磷酸化而被激活,激活后的SnRK2可以作用于下游底物,從而將ABA信號傳遞到不同的信號通路[23]。ABA信號傳遞途徑中,SnRK2在去磷酸化后失活,那么必然存在SnRK2活性的負調控因子,除PP2C外,與SnRK2互作的鈣離子感應因子SCS和一氧化氮(NO)等也是負調控因子[24-25]。SnRK2的下游作用底物有ABA應答元件結合因子ABF、S型陰離子通道SLAC1、K+通道KAT1、NADPH氧化酶AtrdohF等[26-29]。如擬南芥SnRK2.2、SnRK2.3和SnRK2.6通過調控下游AREB/ABF(bZIP類轉錄因子)參與干旱等滲透脅迫的ABA信號轉導途徑[30]。此外,在滲透脅迫條件下,非ABA依賴型SnRK2在未知激酶的作用下發生磷酸化,直接作用于下游基因,進而引發一系列應答反應[31]。
除磷酸化外,SnRK2還通過其他方式調控基因表達,如RNA剪接、mRNA降解、microRNA積累、DNA甲基化和組蛋白去乙酰化等[32-34]。其中RNA剪接是ABA應答反應的一種重要調控機制。基于磷酸化蛋白組學的方法,鑒定了6種與RNA剪接相關的功能蛋白,包括已經證實的RNA剪接因子BTR1L(AtSnRK2.6的底物)[32]。組蛋白去乙酰化介導的表觀修飾也參與基因轉錄調控。SNL2是組蛋白去乙酰化復合物的重要組成成分,SnRK2可能通過磷酸化SNL2調節組蛋白去乙酰化[32]。可見,SnRK2在轉錄、翻譯、表觀修飾等多個層面參與調控基因表達。
3 SnRK2的功能
3.1 SnRK2參與響應逆境脅迫應答
SnRK2參與響應激素、鹽、旱、極端溫度、養分、病害等各種非生物和生物逆境脅迫。SnRK2基因啟動子區通常含有與激素、逆境脅迫應答相關的順式作用元件,如ABA應答元件ABRE、赤霉素應答元件GARE-motif、茉莉酸甲酯應答元件CGTCA-motif、熱激應答元件HSE、低溫脅迫應答元件LTRE、干旱應答元件DRE、逆境脅迫響應元件TC-rich repeats、MYB轉錄因子結合元件MBS等,這些元件可能與激素信號轉導和逆境脅迫應答有關[35]。
第一個被克隆的SnRK2成員是PKABA1,它是從ABA處理后的小麥幼胚cDNA文庫中分離獲得[26]。PKABA1可以磷酸化底物TaABF(ABA應答元件結合因子),并受ABA、冷害、高鹽、干旱等誘導表達[26]。
在擬南芥中,除AtSnRK2.9外,其余9個SnRK2成員均可被蔗糖、甘露醇、山梨醇和NaCl誘導。其中,AtSnRK2.2、AtSnRK2.3、AtSnRK2.6、AtSnRK2.7和AtSnRK2.8(Group II和III)均能夠不同程度地被ABA誘導,但所有AtSnRK2成員都不受低溫誘導[5]。
水稻的10個SnRK2成員均能被鹽脅迫誘導,然而僅有SAPK8、SAPK9和SAPK10這3個成員可以被ABA激活[11]。玉米的11個SnRK2成員中,ZmSnRK2.2、ZmSnRK2.4、ZmSnRK2.5、ZmSnRK2.7和ZmSnRK2.10能被ABA誘導激活,ZmSnRK2.3和ZmSnRK2.6能夠被鹽脅迫強烈誘導,ZmSnRK2.3和ZmSnRK2.7能被低溫誘導,ZmSnRK2.5、ZmSnRK2.6和ZmSnRK2.9的表達會被高溫抑制[17]。
過表達玉米SAPK8的擬南芥[36]和過表達水稻SAPK4的水稻[37],耐鹽性較野生型增強。NaCl溶液處理后,水稻SAPK4基因通過降低氧化損傷和調節離子平衡,細胞中積累較少的Na+和Cl-,從而提高了轉基因植株的耐鹽性[37]。過表達小麥TaSnRK2.4基因會誘導下游抗逆相關基因的表達,引起一系列抗逆生理變化,如組織的滲透勢降低、相對含水量增加、細胞膜穩定性增強等,從而提高了轉基因植株的耐鹽性,還增強了其對低溫、干旱的抗性[38]。
SnRK2也可以參與養分脅迫應答。萊茵衣藻中,PSR1轉錄因子涉及磷和硫的代謝,其中SNRK2.1和 SNRK2.2響應低硫脅迫[39]。高等植物中,在硫不足時擬南芥AtSnRK2.3調控硫酸轉運蛋白SULTR2-2的表達和O-乙酰基-L-絲氨酸(硫饑餓反應推定的信號化合物)的積累[40]。小麥TaSnRK2.7-B和控制磷素高效利用的位點均位于染色體2AL[41]。利用篩選酵母文庫獲得小麥TaSnRK2.4候選互作蛋白TaPHR1(磷素應答蛋白),經酵母雙雜、熒光素酶互補成像和熒光定量PCR技術驗證,推測TaSnRK2.4可能通過磷酸化TaPHR1參與磷素的吸收利用(未發表數據)。除了非生物脅迫外,SnRK2還參與響應病原菌等引起的生物脅迫應答,在植物與病原物互作初期的信號識別與轉導,以及后期的防御反應調控等階段發揮重要作用。SnRK2還可以通過調控植物的氣孔關閉來避免病菌的侵害,如擬南芥AtSnRK2.6基因可能通過調控OST1(open stomata 1)基因表達參與氣孔開閉過程[42]。擬南芥受到無毒丁香假單胞番茄變種Pst DC3000/avrRpt2侵染后,葉片中SnRK2.8的轉錄水平明顯上升,且SnRK2.8能與植物系統抗病的關鍵調控轉錄因子NPR1互作。磷酸化試驗表明,SnRK2.8可以磷酸化NPR1的589位Ser和373位Thr[43]。
3.2 SnRK2參與調控植物生長發育
SnRK2調控種子休眠與萌發、性別分化、根系形態建成、開花、果實成熟、產量形成、株高等多個環節,從而影響植物的生長發育進程[32,44-50]。以下集中介紹SnRK2在調節植物根系生長、果實成熟和農藝性狀發育三個方面的研究進展。
根系是植物吸收水分和養分的重要器官,在穩固植株、營養儲存、繁殖、次級代謝物合成等方面具有重要作用。研究表明,擬南芥AtSnRK2.4和AtSnRK2.10分別在維持根長及側根數量方面發揮重要作用[20],并參與調控鹽脅迫下根系的生長與形態建成[51];干旱脅迫下擬南芥AtSnRK2.8(SRK2C)的突變導致了側根數量減少、根長變短[46];AtSnRK2.2則調控根系的向水性生長[52]。小麥TaSnRK2.3、TaSnRK2.4、TaSnRK2.7和TaSnRK2.8過量表達均能促進轉基因擬南芥根系的生長[38,53-55]。SnRK2參與根系生長的機理值得深入研究。
果實成熟是一個復雜的生理過程。在櫻桃果實成熟過程中,PacSnRK2.1、PacSnRK2.3、PacSnRK2.4和PacSnRK2.6基因均呈現出“上調-下調-上調”的規律性表達模式[12]。蘋果MdSnRK2.4/2.9可與調節乙烯合成酶ACO1表達的兩個關鍵轉錄因子 MdHB1和MdHB2互作,并通過磷酸化協同調控MdHB1和MdHB2的轉錄活性及穩定性,從而調控乙烯合成,影響果實成熟[56]。
SnRK2參與糖代謝,影響產量。作為主要碳源,糖不僅是能量代謝的物質基礎和細胞的組成成分,還是重要的滲透調節物質和信號分子。有學者推測SnRK2和SnRK3均由SnRK1復制演化而來,所以保留了SnRK1的糖代謝功能,與產量密切相關[49],如擬南芥AtSnRK2.6參與調控了蔗糖代謝和不飽和脂肪酸的合成[57]。小麥TaSnRK2.8-A與灌漿中期和成熟期的莖稈可溶性糖含量顯著關聯[58],不同水分條件下TaSnRK2.10-A與千粒重、單株穗數和每穗小穗數等產量性狀顯著關聯[59]。多種環境條件下TaSnRK2.9-5A與穗粒數、千粒重顯著關聯,過表達TaSnRK2.9-5A能顯著增加轉基因水稻的穗粒數[60]。SnRK2增產的分子機理尚未闡明。
SnRK2參與調控株高。關聯分析結果表明,小麥TaSnRK2.3-1B與株高、穗下節長、莖稈倒二節長和千粒重顯著關聯[50]。借助DH群體將TaSnRK2.3-1B定位于小麥染色體1B的標記 wmc156(2.1 cM)和P3446-183(2.9 cM)之間,在該區域內曾報道有一個控制株高的QTL[53]。對小麥極端株高品種的節間組織進行基因表達量分析的結果表明,TaSnRK2.3-1B負調控株高[61]。
綜上所述,SnRK2是典型的“一因多效”基因家族,它一方面響應逆境脅迫應答,增強植物的抗逆性,一方面參與調控植物生長發育的多個方面。
4 展 望
SnRK2是植物特有的蛋白激酶,它通過磷酸化修飾調節蛋白質活性,調控下游基因表達,從而參與多種信號轉導途徑和代謝過程,在植物生長發育和逆境脅迫應答中發揮重要作用。
目前對SnRK2的研究主要集中在ABA依賴的信號通路,其作用機理和分子調控網絡也因此被闡述得很清楚。而對SnRK2參與的非ABA依賴信號途徑研究得卻非常之少,其上游激活因子、下游靶標,以及其作用機理均知之甚少,因此未來應該加強對該領域的研究。
隨著轉基因、高通量測序和基因組編輯技術的廣泛應用,基因功能解析將變得越來越容易;同時伴隨著關聯分析技術在復雜性狀解析方面的成功應用,我們深信SnRK2的新功能將會被不斷發現,尤其是其在調控主要農藝性狀發育方面的作用將進一步豐富我們的認知。SnRK2功能的全面解析,將為充分利用該基因家族改良植物的抗逆性、提高產量提供理論依據和基因資源。
猜你喜歡
少兒科學周刊·兒童版(2021年19期)2021-12-10 14:13:40
小學閱讀指南·低年級版(2021年3期)2021-03-19 06:12:40
小天使·二年級語數英綜合(2020年8期)2020-12-23 04:57:40
小天使·一年級語數英綜合(2020年11期)2020-12-16 02:57:22
學苑創造·A版(2020年3期)2020-04-24 09:21:39
小溪流(畫刊)(2017年11期)2018-01-09 19:15:14
少兒科學周刊·兒童版(2017年5期)2017-06-29 22:24:28
少兒科學周刊·兒童版(2017年5期)2017-06-29 16:46:33
紅領巾·萌芽(2017年5期)2017-06-23 10:35:59
爆笑show(2016年7期)2017-02-09 09:36:13
