脂滴在病原微生物感染中作用的研究進展
2019-08-07 04:55:38嚴夢楠綜述李華茵審校
復旦學報(醫學版) 2019年4期
嚴夢楠(綜述) 周 建 李華茵(審校)
(復旦大學附屬中山醫院呼吸內科 上海 200032)
脂滴是在19世紀由科學通過光學顯微鏡發現[1]。在20世紀90年代以前,認為脂滴在細胞中不發揮任何功能而忽視了對它的研究。直到1991年發現perilipin (PLIN)蛋白特異定位在脂滴表面,才開始關注脂滴的生物學功能[2]。脂滴在脂質代謝和維持機體能量穩態等方面起到重要的作用,脂滴功能受損會導致多種疾病。病原微生物可以利用脂滴進行生命活動,完成自身復制及其在宿主中的持續存在,而脂滴也可在一定程度上起到抵御病原微生物入侵細胞的作用。本文對近年來脂滴在病原微生物感染中作用的研究進展作一綜述。
脂滴的概述脂滴廣泛分布于真核細胞,在一些原核生物中有存在,是細胞內中性脂的儲存場所[3-4]。
脂滴的形成 因為合成中性脂的酶類主要分布在內質網,因此目前普遍認為脂滴是在內質網形成的。雖然有不同的模型解釋從中性脂合成到脂滴形成的過程,但均未被完全證實。較廣泛接受的模型是中性脂在內質網雙層膜中積聚并形成“透鏡樣脂質結構(lipid lens)”,積聚至一定程度后從內質網以出芽(budding)的方式形成新生脂滴并繼續生長,最終形成成熟脂滴[5-6]。
脂滴的生物學特征及功能 脂滴由極性單磷脂層包裹疏水核心組成,是細胞內諸如三酰甘油、膽固醇酯等中性脂的貯存場所[7-8]。在脂肪細胞中,脂滴的直徑可達到100 μm,而脂滴在非脂肪細胞中的直徑則可小于1 μm[4]。一直以來,人們認為脂滴的主要功能是儲存能量。隨著蛋白質組學技術的發展與應用,在脂滴表面發現了許多功能蛋白,如PLIN蛋白家族中的PLIN1蛋白、PLIN2蛋白[又稱為脂肪分化相關蛋白(adipose differentiation-related protein,ADRP)]以及PLIN3蛋白[又稱為47 000的尾連蛋白(tail-interacting protein of 47 000,TIP47)][9]。同時發現脂滴能夠與其他細胞結構如內質網、線粒體、過氧化物酶體、核內體等相互作用,以維持細胞穩態和應對不利情況[10-11]。脂滴的功能不僅局限于脂類儲存與代謝。脂滴通過將游離脂肪酸轉化為中性脂,可以有效避免積聚的脂肪酸及其衍生物對細胞的毒性作用[6]。脂滴在維持內質網穩態中也發揮了重要作用。當體內環境改變或有刺激因素存在時,會啟動內質網應激(endoplasmic reticulum stress,ERS),并引起未折疊蛋白反應(unfold protein response,UPR),與此同時,脂滴也會相應增加,從而有助于清除錯誤折疊蛋白并恢復內質網脂質代謝平衡[12]。脂滴在自噬中也發揮了一定的調節作用[13]。當細胞營養不足時會引起自噬水平上調,導致氨基酸及脂類釋放增多,而釋放的脂類再被酯化成三酰甘油后進入脂滴。這一過程可以減少脂毒性對線粒體的損傷,保證線粒體功能正常[6]。此外,脂滴在膜轉運和合成、蛋白質貯存和降解、信號傳導、炎癥反應等過程中也發揮重要作用。因此,脂滴被認為是一個復雜且處于動態變化的細胞器[8,11,14-15]。
脂滴在病原微生物感染中的作用近年來研究發現,包括病毒、細菌、寄生蟲在內的一些病原微生物都能與脂滴相互作用。最初認為病原微生物主要是利用脂滴來獲得能量及脂質等來源,而研究證據顯示脂滴也可以在免疫反應中發揮媒介作用。
為病原微生物提供營養物質及能量 結核分枝桿菌感染宿主肺泡巨噬細胞后,局部炎癥反應導致免疫細胞向感染部位聚集并形成肉芽腫。肉芽腫的中心由泡沫巨噬細胞和朗格漢斯多核巨細胞包圍感染的巨噬細胞,周圍由淋巴細胞、成纖維細胞環繞。泡沫巨噬細胞的形成是分枝桿菌感染的典型特征之一[16]。研究發現,包括結核分枝桿菌(Mycobacteriumtuberculosis)、牛型分枝桿菌(Mycobacteriumbovis)、麻風桿菌(Mycobacteriumleprae)、海魚分枝桿菌(Mycobacteriummarinum)等在內的細菌感染宿主細胞后,均會導致脂滴重新分布,脂滴被認為是分枝桿菌慢性感染中主要的能量和碳來源[9]。海魚分枝桿菌感染宿主細胞后,宿主細胞內原先散在分布的脂滴就會向吞噬了細菌的吞噬體處聚集,用熒光染料Bodipy493/503標記脂滴,吞噬體內中性脂的熒光強度增強。將熒光染料Bodipy558/568 C12標記的脂肪酸與宿主細胞共同培養,使宿主合成的脂質帶熒光標記,同時細菌體內也出現熒光標記[17](圖1)。結核分枝桿菌中有脂肪代謝相關酶類,編碼三酰甘油合酶-1 (triacylglycerol synthase 1,Tgs1)的基因,可利用宿主來源的脂肪酸合成三酰甘油,并存儲在細菌自身的脂滴中。結核分枝桿菌還編碼脂肪酶LipY,可分解細菌儲存的脂肪以供能,也可分解宿主細胞的三酰甘油來供細菌利用[18]。這些研究認為在分枝桿菌感染過程中,脂滴起到供能及為細菌代謝提供所需物質的作用。但Knight等[19]提出與傳統觀念不同的看法,在缺少脂滴的巨噬細胞中,結核分枝桿菌仍可以積累脂質,而巨噬細胞內脂滴形成離不開干擾素-γ(interferon-γ,IFN-γ)的作用。因此認為,結核分枝桿菌感染后,細菌能夠獲取非脂滴來源的營養物質及能量。巨噬細胞內脂滴的形成是依賴IFN-γ的程序化的巨噬細胞免疫反應,主要是為類花生酸的合成提供重要場所。
除了脂質代謝,脂滴還可以將細胞內的鐵通過結核分枝桿菌分泌的親脂性鐵載體運輸給細菌,為細菌提供必要的代謝物質[20]。
在對寄生蟲的研究中同樣發現,克氏錐蟲(Trypanosomacruzi)、剛地弓形蟲(Toxoplasmagondii)、利什曼原蟲(Leishmaniamajor)等寄生蟲也可以誘導宿主細胞中脂滴積聚,同樣導致脂滴的重新分布[21]。這些寄生蟲利用宿主的脂類合成自身的脂滴,完成增殖、復制等生命活動[22-23]。
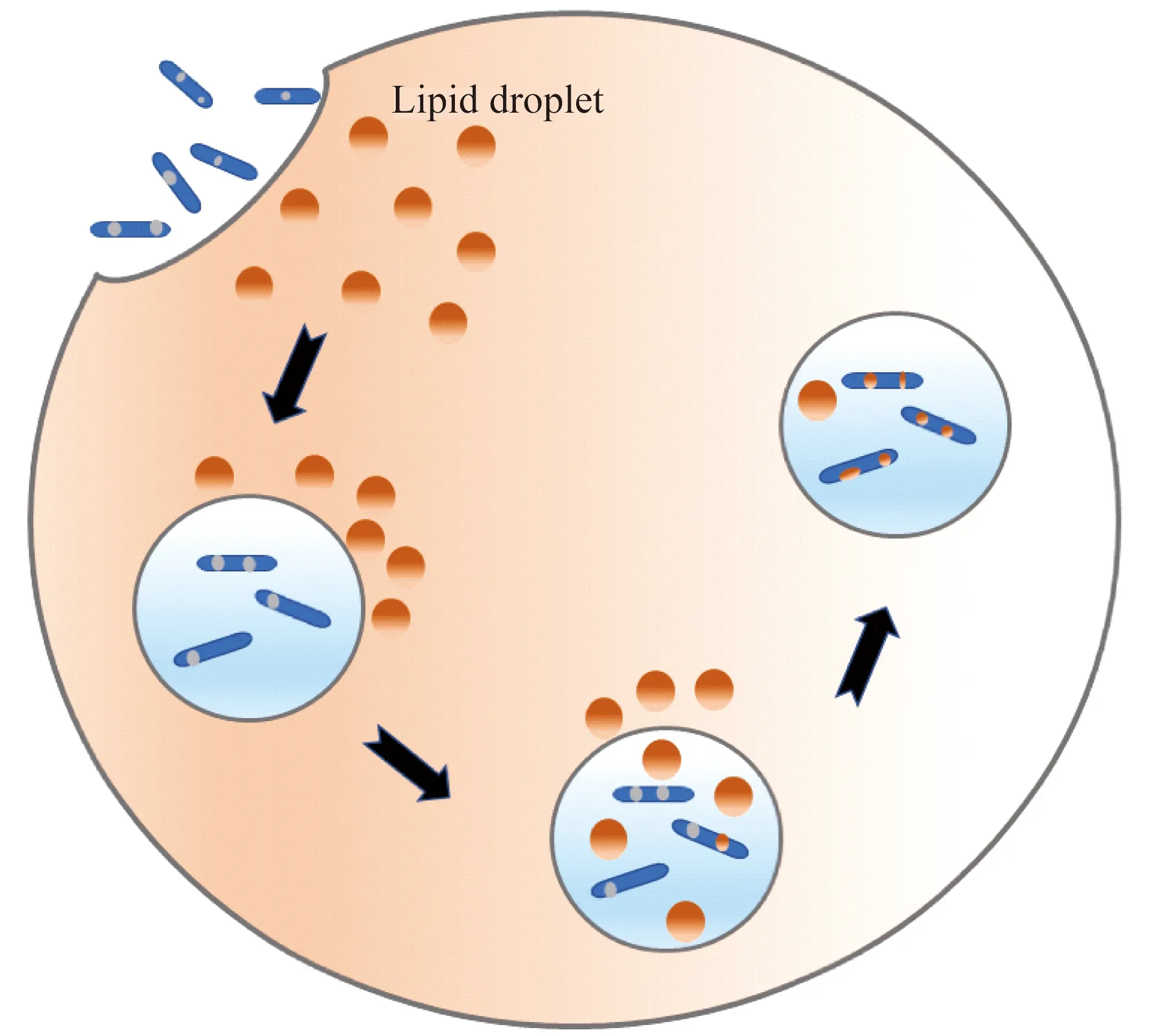
AfterMycobacteriummarinuminfection,LDs relocate to the bacterium-containing compartment and are found to accumulate inside the compartment.Later,LDs are transferred into the bacteria.
圖1 分枝桿菌感染宿主細胞后脂質分布
Fig 1 Distribution of lipids duringMycobacteriummarinuminfection of host cell
為病毒復制提供組裝平臺 丙型肝炎病毒(hepatitis C virus,HCV)的復制發生在內質網上,而組裝則發生在脂滴表面,病毒衣殼蛋白core蛋白和非結構蛋白5 (nonstructural protein 5,NS5A)蛋白在其中發揮重要的作用。HCV感染后,在內質網上形成core蛋白,包括D1和D2兩個結構區域:D1區域為正電區域,可以結合RNA和蛋白質;D2區域則與脂滴結合[24]。NS5A在內質網和脂滴上均有分布[24]。NS5A蛋白與病毒RNA結合,將RNA從復制位點運到脂滴,從而被core蛋白包裹,形成一個完整的病毒顆粒[9]。在組裝過程中,宿主的二酰甘油酰基轉移酶(diacylglycerol acyltransferases 1,DGAT1)在內質網上與core蛋白和NS5A蛋白結合,并促進其正確定位到脂滴表面[25-26]。在組裝過程中,TIP47和Ras相關蛋白18 (Rab18)起到重要作用:TIP47與NS5A相互作用,并依賴TIP47與脂滴的高親和力,將結合了RNA的NS5A運送到脂滴表面[27];Rab18與NS5A結合,使病毒復制復合物與脂滴相互靠近[28](圖2)。與HCV同屬黃病毒科的登革熱病毒(dengue virus,DENV)、GB病毒B(GBV-B)等也都通過core蛋白定位到脂滴表面,完成病毒顆粒組裝[9,15]。

The replication of HCV genome is occurred on the cytoplasmic face of the ER.TIP47 brings RNA-loaded NS5A to the LD for the interaction with HCV core protein,resulting in the encapsidation of the viral RNA.TIP47:Tail-interacting protein of 47 000;Rab18:Ras-related protein 18;NS5A:Nonstructural protein 5 A;DGAT:Diacylglycerol acyltransferases;ER:Endoplasmic reticulum;RNA:Ribonucleic Acid.
圖2 HCV病毒與脂滴相互作用的模式圖
Fig 2 Model of interactions between HCV and lipid droplets
抵御病原微生物的感染 脂滴在病原微生物感染后的免疫反應中發揮重要作用。結核分枝桿菌、HCV、剛地弓形蟲等病原微生物感染都會引起宿主細胞內脂滴數量增加[29-31]。脂滴的核心由磷脂和中性脂組成,在酶的作用下可以釋放花生四烯酸。花生四烯酸是合成前列腺素、白三烯等類花生酸的主要底物,在脂滴中也存在合成類花生酸的酶類,包括環氧化酶(cyclooxygenases,COX)、前列腺素E2合成酶(prostaglandin E2 synthase)、白三烯C4合成酶(leukotriene C4 synthase)等,而類花生酸是參與炎癥反應的重要信號分子[15,32-33]。當宿主細胞遇到病原微生物感染時,脂滴可以針對刺激產生相應的炎癥介質,調節免疫反應。
脂滴上還存在抗病毒的蛋白。病毒抑制蛋白(virus inhibitory protein,Viperin)是由干擾素誘導產生的抗病毒蛋白。Viperin依賴N端的α-螺旋結構將其定位在內質網靠近細胞質的一面以及脂滴表面[34]。Viperin能在脂滴表面通過C末端區域與HCV的NS5A相互作用,從而發揮抑制病毒復制的作用[7]。HCV感染后,core蛋白能夠誘導肝臟的脂肪變性,該作用需要TIP47的參與。敲低TIP47表達后,core蛋白表達相應減少,這可能成為減輕HCV感染后所致病理反應的有效方法[35]。
組蛋白(histone)有抗微生物的作用[36]。游離的組蛋白會對細胞造成嚴重損傷,許多生物都會將細胞質中游離的組蛋白降解掉[37]。從果蠅胚胎中提純的脂滴上發現較多量的組蛋白且在體外實驗中表現出很強的殺菌作用,組蛋白通過Jabba蛋白定位到脂滴上,Jabba突變型果蠅胚胎的脂滴上缺少組蛋白[12]。為了驗證脂滴上的組蛋白在體內是否也能起到保護作用,將大腸埃希菌注入野生型和Jabba突變型果蠅胚胎,發現野生型細菌數量減少,而突變型細菌數量顯著增多。用其他革蘭陽性和革蘭陰性細菌感染這兩型胚胎,野生型存活數較突變型多[7]。因此認為,脂滴能通過組蛋白來保護果蠅免受細菌侵襲。
結語隨著對脂滴的認識不斷加深,其呈現出更多、更復雜的功能。在脂滴與病原微生物相互作用的過程中,仍然有許多問題需要解決,如:分枝桿菌具體是通過什么機制使脂滴得以在宿主細胞中重新分布,以及在脂滴相關研究中抑制劑的應用是否會影響體內脂質穩態,能否保證未來作為治療手段使用時的安全性。通過揭示脂滴與病原微生物相互作用的詳細過程及具體機制,能夠為日后臨床控制并治療病原微生物感染提供新思路和新方法。
