竹節參轉錄組使用密碼子偏好性分析
2019-08-13 08:54:10梁娥齊敏杰丁延慶
江蘇農業科學 2019年2期
梁娥 齊敏杰 丁延慶
摘要:竹節參是我國珍稀瀕危中藥材,研究其基因密碼子使用模式,可為利用基因工程技術實現人參皂苷異源生物合成及竹節參分子育種改良提供理論依據。以竹節參轉錄組測序結果為數據來源,篩選編碼蛋白基因序列(coding sequence,簡稱CDS)堿基數不小于300 bp的 11 199 條完整開放閱讀框序列作為研究對象,利用Codon和SPSS軟件分別統計竹節參基因密碼子GC含量、密碼子第3位的(C+G)含量(GC3)和密碼子第1、第2位(G+C)含量的平均值(GC12)、同義密碼子的相對使用度(RSCU)、有效密碼子數(ENC)等密碼子偏好性指標,通過中性繪圖(GC12 vs. GC3)、PR2繪圖和ENC-GC3s繪圖分析影響竹節參密碼子使用模式的因素。結果表明,竹節參基因的平均GC、GC12和GC3s含量分別為44.67%、46.97%和39.80%,其密碼子使用模式受到突變和選擇等多重因素的影響,確定了31個竹節參最優密碼子,除了UUG外,其余最優密碼子均以A或T結尾。竹節參密碼子使用模式與大腸桿菌和釀酒酵母相比差異較大,選取畢赤酵母作為竹節參基因的異源表達宿主更為合適。
關鍵詞:竹節參;轉錄組;密碼子使用模式;最優密碼子
中圖分類號: Q755;S567.5+10.1 ?文獻標志碼: A ?文章編號:1002-1302(2019)02-0059-05
遺傳密碼子是生物體DNA與蛋白質之間信息傳遞的基本單位,具有簡并性,即同一氨基酸有多個對應的密碼子,編碼同一種氨基酸的密碼子叫作同義密碼子,同義密碼子在同一物種不同基因間或不同物種內的使用頻率大有不同,這種不均衡使用模式稱為密碼子使用偏好性,通常把使用頻率較高的一種或幾種同義密碼子稱為最優密碼子[1-2]。研究顯示,不同物種之間基因密碼子偏好性是由突變壓力(如GC含量、基因堿基組成)和自然選擇作用(如翻譯起始信號、基因表達水平、蛋白結構與長度、tRNA豐度等)引起的,mRNA的二級結構及其穩定性、翻譯的速度和準確度、蛋白質折疊等因素也與密碼子的偏好性有關[3-4]。對物種密碼子偏好性開展研究,有助于理解物種進化發展及密碼子使用偏好性的調控機制。密碼子偏好性在基因異源表達研究方面也顯示了重要作用,基因的表達量越大,其密碼子偏好性越強[5],根據這一原理,替換基因低表達密碼子可以提高外源基因表達量,同時根據密碼子使用偏好性可以選擇更為合適的宿主表達系統。有報道表明,可通過優化密碼子的方式來提高外源基因在宿主細胞中的表達量[6-7]。周宗梁等通過優化密碼子的方法提高了蘇云金芽孢桿菌基因cry1Ah在玉米和水稻中的表達量[8-9];楊金玲等通過優化蝎毒鎮痛活性肽基因BmKAngM1,將其導入畢赤酵母后該基因表達量得到顯著提高[10];通過使用最優密碼子,在草菇[11]、擬南芥[12]、川母貝[13]、菠蘿[14]等生物中均得到了很好的研究成果。
竹節參(Panax japonicus C. A. Mey)為多年生草本植物,屬于五加科(Paeoniaceae)人參屬,是我國珍稀瀕危的“七類中草藥”之一,具有抗炎、延緩衰老、降血糖等藥理作用,有著極高的藥用和保健價值。竹節參中富含活性物質三萜皂苷,也是其特征性成分。目前在竹節參種質資源[15-16]、毛狀根的培養[17]、基因工程代謝的調控[18]、生藥學鑒定[19]、精油成分分析[20]和三萜皂苷代謝追蹤[21]等幾個研究領域已經開展了許多研究工作,但直接從竹節參中通過分離提取的方法獲得三萜皂苷對資源消耗極大,技術難度較高。若利用生物合成的方法則可以很好地解決這一困難,實現有效成分的生物合成,選擇適合關鍵酶基因高效表達的異源表達系統是一個重要步驟。本研究以竹節參轉錄組數據為材料,通過分析竹節參基因密碼子組成的各項指標,研究竹節參表達基因密碼子使用偏好性及其影響因素,以期為竹節參相關基因表達系統的選擇及分子育種提供理論基礎。
1 材料與方法
1.1 數據來源
竹節參轉錄組數據來源于文獻[22],通過Perl語言程序對竹節參轉錄組數據進行過濾篩選,篩選出堿基數≥300 bp的蛋白質編碼序列共11 199條,作為密碼子分析的數據來源。本研究中使用到的大腸桿菌(Escherichia coli)、釀酒酵母(Saccharomyces cerevisiae)和畢赤酵母(Pichia pastoris)的密碼子偏好性數據來自Codon Usage database( http://www. kazusa. or. jp /codon /)。
1.2 竹節參基因GC含量分析及中性繪圖
利用Codon W1.4.2(http://codonw.sourceforge.net/)統計分析竹節參基因密碼子的堿基組成規律,測得鳥嘌呤和胞嘧啶總體含量G+C、密碼子第3位堿基組成(A3、G3、C3、T3)、密碼子第3位的C+G數(GC3)和密碼子第1、第2位的平均G+C含量(GC12)等數據。本研究以密碼子第1、第2個堿基的G+C平均含量為縱坐標,以密碼子第3個堿基的G+C含量為橫坐標,通過中性繪圖探究影響密碼子偏好性的因素。分析密碼子第1、第2位與第3位堿基組成的相關性,來分析自然選擇與壓力突變對基因密碼子使用模式的影響[23]。
1.3 竹節參基因有效密碼子數分析
參照Fuglsang的方法[24],利用有效密碼子數來衡量竹節參基因單個密碼子使用的偏好程度。基因有效密碼子數(effective number codon,簡稱ENC)是衡量基因密碼子偏好性的一個重要指標,數值范圍為20(每個氨基酸只使用1個同義密碼子的極端偏好情況)到61(每個同義密碼子被平均使用的無偏好情況)。研究表明,當ENC值≤35時,基因密碼子的使用偏好性越強[25]。同時以ENC值為縱坐標,GC3s為橫坐標進行ENC繪圖,分析各基因密碼子的使用特征,研究竹節參密碼子偏好性和基因堿基組成之間的關系。
1.4 同義密碼子相對使用度及最優密碼子分析
參照Sharp等的方法[26],采用同義密碼子的相對使用度(RSCU)作為衡量竹節參密碼子使用偏好性的指標。使用3個指標表示:密碼子使用無偏好性(RSCU=1);密碼子使用度較大(RSCU>1);密碼子使用頻率較低(RSCU<1)。利用高表達優越密碼子分析方法[27],統計所有基因的ENC值、有序數據集的上下10%區間形成高RSCU集合和低RSCU集合,進行最優密碼子分析。根據2個子集的ΔRSCU值及卡方檢驗確定最優密碼子。
1.5 PR2繪圖分析
采用PR2繪圖進行密碼子奇偶偏好分析[23],分別統計竹節參基因中A3/(A3+T3)與G3/(G3+C3)數據,以此為縱坐標和橫坐標,通過平面圖顯示各基因堿基的組成。為使編碼密碼子第3位堿基對A/T或T/A和G/C或C/G的突變不均衡,PR2繪圖時只選擇亮氨酸(Leu,密碼子:CTA、CTC、CTG、CTT)、脯氨酸(Pro)、精氨酸(Arg,密碼子:CGA、CGC、CGG、CGT)、絲氨酸(Ser,密碼子:TCA、TCC、TCG、TCT)、丙氨酸(Ala)、纈氨酸(Val)、蘇氨酸(Thr)和甘氨酸(Gly)等遺傳密碼子進行分析。
2 結果與分析
2.1 GC含量分析以及中性繪圖
竹節參轉錄組數據中共有11 199條完整的開放閱讀框序列,利用Codon W對完整的開放閱讀框序列進行密碼子使用模式分析。結果(表1)表明,所有完整的開放閱讀框序列總長度為 12 006 732 bp,N50=1 332 bp,平均GC含量為44.67%,GC含量范圍為30.3%~63.8%。竹節參基因組平均GC含量低于大腸桿菌基因組平均GC含量(52.35%),高于釀酒酵母基因組平均GC含量(39.77%)和畢赤酵母基因組平均GC含量(42.73%)。竹節參基因組密碼子第1、第2位堿基GC含量的變化范圍為31.7%~71.8%,平均GC含量為46.97%。密碼子的第3位上堿基GC含量的變化范圍為10.7%~83.9%,平均GC含量是39.80%。密碼子第3位堿基上A、T的使用頻率分別是33.09%、41.27%,略高于C、G的使用頻率(分別為23.61%、27.57%),說明竹節參基因密碼子對以A和T結尾的密碼子的使用偏好程度較大,而對以G和C結尾的密碼子的使用偏好程度較小。密碼子第3位堿基上的平均GC含量(39.80%)略低于畢赤酵母基因密碼子第3位堿基的平均GC含量(42.16%),偏高于釀酒酵母密碼子第3位堿基的平均GC含量(38.10%),大大低于大腸桿菌遺傳密碼子第3位堿基的GC平均含量(55.62%)。研究表明,竹節參對密碼子使用并無特殊堿基偏好,其密碼子使用模式與大腸桿菌和釀酒酵母差異較大,與畢赤酵母差異略小。如圖1所示,中性繪圖分析表明,竹節參密碼子GC12取值范圍為31.7%~71.8%,GC3的取值范圍為10.7%~83.9%,GC3s與GC12的相關系數為0.062,回歸系數為0.48,表明并無顯著相關性,說明竹節參基因密碼子使用模式主要受到自然選擇的影響。
2.2 竹節參基因有效密碼子數分析
研究顯示,竹節參ENC值一般在25.88~61.00,ENC平均值為53.04。通常將ENC值=35作為衡量密碼子偏好性強弱的標準[22]。竹節參基因有13條ENC值小于35,表明竹節參基因整體水平密碼子偏好性較低,只有少數基因具有密碼子偏好性。由表2的ENC相關性分析可知,GC與GC12、GC3之間的相關性均達極顯著水平,GC12和GC3之間并沒有明顯的相關性,密碼子成分不相同,而ENC值與密碼子數的相關性也沒有達到顯著水平,說明密碼子數對ENC的影響很弱,排除了基因長度過短對密碼子偏好性的影響。如圖2所示,ENC-GC3s繪圖分析結果表明大部分竹節參基因均分布在標準曲線的周圍,而小部分基因則分布在距標準曲線較遠的位置,說明竹節參表達基因密碼子使用模式不僅受到自然選擇的影響,同時也受到突變壓力作用的影響。
2.3 同義密碼子的相對使用度及最優密碼子分析
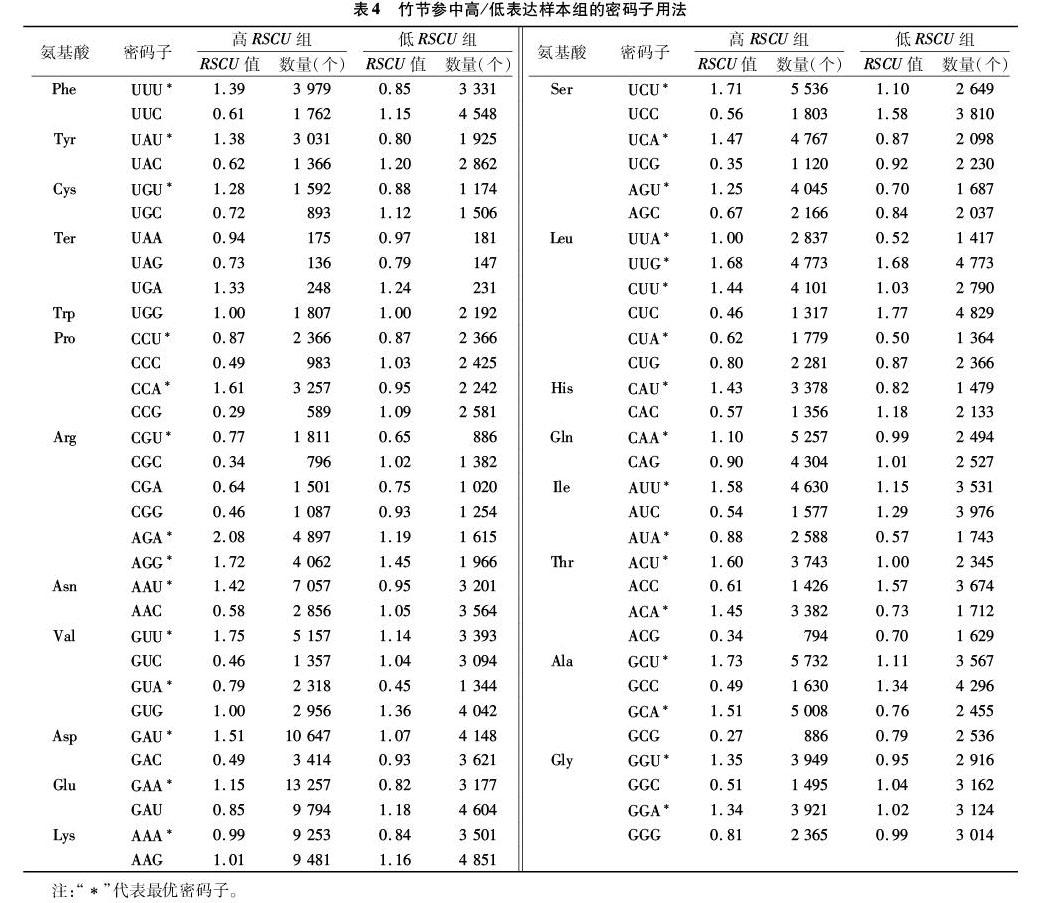
如表3所示,竹節參共有28個密碼子的RSCU值大于1,說明這28個密碼子可能是竹節參偏好使用的。除去Trp和Met(只有1種密碼子編碼)外,編碼Phe、Ser、Leu、Tyr、Cys、His等氨基酸的密碼子及終止密碼子存在較大的偏好性,RSCU>1的密碼子主要以U和A結尾。如表4所示,對竹節參基因進行高、低表達樣本庫比較,篩選出竹節參轉錄組樣本的最優密碼子共31個,分別是UUU、UUA、UUG、CUU、CUA、AUU、AUA、GUU、GUA、UAU、CAU、CAA、AAU、AAA、GAU、GAA、UCU、UCA、CCU、CCA、ACU、ACA、GCU、GCA、UGU、CGU、AGU、AGA、AGG、GGU、GGA,它們分別編碼苯丙氨酸、亮氨酸、異亮氨酸、纈氨酸、酪氨酸、組氨酸、谷氨酰胺、天冬酰胺、賴氨酸、天冬氨酸、谷氨酸、絲氨酸、脯氨酸、蘇氨酸、丙氨酸、半胱氨酸、精氨酸和甘氨酸共18個氨基酸,其中Arg有4個最優密碼子。在這31個最優密碼子中,除了UUG是以G結尾外,其他密碼子均是以A或T結尾的,說明竹節參偏好使用以A/T結尾的密碼子。
2.4 PR2-plot分析
通過PR2-plot分析,表明了竹節參各基因密碼子中4個堿基組分嘌呤(A和G)與嘧啶(T和C)之間的關系,如圖3所示,4個堿基均不均衡使用,竹節參密碼子第3位堿基T的使用頻率大于堿基A,堿基G的使用頻率大于堿基C,表明竹節參基因密碼子使用模式受到突變壓力和自然選擇等多重因素的影響。
3 討論
每種生物的特有密碼子使用模式是物種為適應自然環境而進化形成的。作為生物基因組中堿基組成的一項重要指標,GC含量的高低表明了生物突變方向性的頻數大小。研究表明,密碼子第3位上GC堿基的含量受到突變壓力的影響較小,因此GC3s含量通常被作為分析密碼子使用模式的一個主要參數。本研究分析竹節參基因GC含量分布情況,發現平均GC含量和GC12比較接近(均低于50%),表明所有竹節參基因中整體AU含量略高于GC,且密碼子主要以A和U結尾。相比于畢赤酵母,其密碼子使用模式與大腸桿菌和釀酒酵母的差異更大。多數研究者對高等植物如水稻(Oryza sativa)、小麥(Triticum aestivum)、大麥(Hordeum vulgare)、真菌與一些單細胞微生物如鐮狀瘧原蟲(Plasmodium falciparum)、支原體(Mycoplasmacapricolum)和植物線粒體微基因組中密碼子的使用模式進行了研究,結果表明高等植物基因密碼子中GC含量相對較高且趨于以G/C結尾[25],在低等生物中則AU含量明顯高于GC含量[26-31]。研究表明,物種密碼子使用模式在形成過程中往往受到很多因素的影響,如突變壓力和自然選擇等。本研究中中性繪圖分析結果表明,GC12與GC3s二者之間其實不存在明顯的相關性;ENC-plot分析表明,竹節參基因大部分均分布在標準曲線周圍,而小部分基因則分布于距標準曲線較遠的位置;PR2-plot 分析發現,竹節密碼子A、C、T和G 4個堿基使用不均衡,結合中性繪圖、ENC-plot和 PR2-plot綜合分析表明,竹節參基因密碼子使用模式受到突變壓力和自然選擇等多重因素的影響。通過高偏性與低偏性基因庫RSCU值的比較,利用ΔRSCU大小等級劃分以確定最優密碼子[32-35],本研究中通過此方法篩選出竹節參基因偏好密碼子共31個,在31個密碼子中,除了UUG外,其余最優密碼子均以A或T結尾,以此推測竹節參基因密碼子的偏好性與GC3s含量呈負相關。有研究者對長春花[36]、小麥[23]等植物密碼子使用模式進行研究發現,最優密碼子大多是以A/T結尾的,且通常表現出對嘌呤堿基U的偏好強于嘌呤堿基A,與本研究結果相似。本研究以竹節參轉錄組為數據來源,研究其密碼子使用模式,確定竹節參表達基因的最優密碼子,一方面可以利用密碼子優化對外源基因進行密碼子改造,提高外源基因在竹節參中的表達水平,同時也為竹節參相關基因的異源表達及分子育種等提供了理論依據。
參考文獻:
[1]Ikemura T. Codon usage and tRNA content in unicellular and multicellular organisms[J]. Molecular Biology and Evolution,1985,2(1):13-34.
[2]朱孝軒,朱英杰,宋經元,等. 基于全基因組和轉錄組分析的赤芝密碼子使用偏好性比較研究[J]. 藥學學報,2014(9):1340-1345.
[3]Fedorov A,Saxonov S,Gilbert W. Regularities of context-dependent codon bias in eukaryotic genes[J]. Nucleic Acids Research,2002,30(5):1192-1197.
[4]Hiraoka Y,Kawamata K,Haraguchi T A. Codon usage bias is correlated with gene expression levels in the fission yeast Schizosaccharomyces pombe[J]. Genes to Cells,2009,14(4):499-509.
[5]Quax T E,Claassens N J,Soell D,et al. Codon bias as a means to fine-tune gene expression[J]. Molecular Cell,2015,59(2):149-161.
[6]Zelasko S,Palaria A,Das A. Optimizations to achieve high-level expression of cytochrome P450 proteins using Escherichia coli expression systems[J]. Protein Expression and Purification,2013,92(1):77-87.
[7]李麗莎,李祥龍,周榮艷,等. 山羊酪氨酸相關蛋白1(TYRP1)基因密碼子偏好性分析[J]. 貴州農業科學,2016,44(3):113-119.
[8]周宗梁,林智敏,耿麗麗,等. 水稻中cry1Ah1基因密碼子優化方案的比較[J]. 生物工程學報,2012,28(10):1184-1194.
[9]李圣彥,郎志宏,朱 莉,等. 利用密碼子優化提高Bt cry1Ah基因在轉基因玉米(Zea mays L.)中的表達[J]. 中國農業科技導報,2011,13(6):20-26.
[10]楊金玲,高麗麗,朱 平,等. 蝎毒鎮痛活性肽基因BmK AngM1的密碼子優化及其真核表達分析[J]. 藥學學報,2012(10):1389-1393.
[11]蔣 瑋,呂貝貝,何建華,等. 草菇密碼子偏好性分析[J]. 生物工程學報,2014,30(9):1424-1435.
[12]范三紅,郭藹光,單麗偉,等. 擬南芥基因密碼子偏愛性分析[J]. 生物化學與生物物理進展,2003,30(2):221-225.
[13]李 瀅,匡雪君,孫 超,等. 川貝母轉錄組密碼子使用偏好性分析[J]. 中國中藥雜志,2016,41(11):2055-2060.
[14]陳 哲,胡福初,王祥和,等. 菠蘿轉錄組基因密碼子使用偏好性分析[C]//中國熱帶作物學會2016年學術年會論文集,2016:2.
[15]張 來. 黔產竹節參種子萌發試驗研究[J]. 種子,2012,31(5):75-78.
[16]羅正偉,張 來,呂翠萍,等. 竹節參離體培養及植株再生[J]. 中藥材,2011,34(12):1818-1823.
[17]張 來,張顯強,羅正偉,等. 竹節參毛狀根培養體系的建立及人參皂苷Re的合成[J]. 中國中藥雜志,2010,35(18):2383-2387.
[18]Zhang L,Sun M. Molecular cloning and sequences analysis of SS gene from Panax japonicus[J]. Research Journal of Biotechnology,2014,9(6):59-63.
[19]張 來,孫 敏. 貴州民間苗藥竹節參葉的生藥鑒定[J]. 中藥材,2009,32(5):691-693.
[20]Meyer C A,Zhang L,Zhang X Q,et al. Comparative analysis of the essential oils from normal and hairy roots of Panax japonicas[J]. African Journal of Biotechnology,2011,10(13):497-519.
[21]張 來,楊碧昌,黃元射. 黔產竹節參根人參皂苷提取工藝與RP-HPLC含量分析[J]. 廣東農業科學,2015,42(20):86-90.
[22]Rai A,Yamazaki M,Takahashi H,et al. RNA-seq transcriptome analysis of panax japonicus,and its comparison with other panax species to identify potential genes involved in the saponins biosynthesis[J]. Frontiers in Plant Science,2016,7(e0144):481.
[23]Zhang W J,Zhou J,Li Z F,et al. Comparative analysis of codon usage patterns among mitochondrion,chloroplast and nuclear genes in Triticum aestivum L.[J]. Journal of Integrative Plant Biology,2007,49(2):246-254.
[24]Fuglsang A. The effective number of codons for individual amino acids:some codons are more optimal than others[J]. Gene,2003,320(3):185-190.
[25]Comeron J M,Aguadé M. An evaluation of measures of synonymous codon usage bias[J]. Journal of Molecular Evolution,1998,47(3):268-274.
[26]Sharp P M,Li W H. The codon adaptation index—a measure of directional synonymous codon usage bias,and its potential applications[J]. Nucleic Acids Research,1987,15(3):1281-1295.
[27]Bellgard M,Schibeci D,Trifonov E,et al. Early detection of G+C differences in bacterial species inferred from the comparative analysis of the two completely sequenced helicobacter pylori,strains[J]. Journal of Molecular Evolution,2001,53(4/5):465.
[28]Kawabe A,Miyashita N T. Patterns of codon usage bias in three dicot and four monocot plant species[J]. Genes & Genetic Systems,2003,78(5):343.
[29]Saul A,Battistutta D. Codon usage in Plasmodium falciparum[J]. Molecular and Biochemical Parasitology,1988,27(1):35-42.
[30]Muto A,Yamao F,Osawa S. The genome of Mycoplasma capricolum[J]. Progress in Nucleic Acid Research & Molecular Biology,1987,34:29.
[31]Dybvig K,Voelker L L. Molecular biology of Mycoplasmas[J]. Annual Review of Microbiology,2003,50(50):25-57.
[32]劉慶坡,薛慶中. 粳稻葉綠體基因組的密碼子用法[J]. 作物學報,2004,30(12):1220-1224.
[33]續 晨,賁愛玲,蔡曉寧. 蝴蝶蘭葉綠體基因組密碼子使用的相關分析[J]. 分子植物育種,2010,8(5):945-950.
[34]尚明照,劉 方,華金平,等. 陸地棉葉綠體基因組密碼子使用偏性的分析[J]. 中國農業科學,2011,44(2):245-253.
[35]李秀璋,宋 輝,李春杰. 茄腐鐮孢(Fusarium solani)線粒體基因組密碼子偏好性分析[J]. 基因組學與應用生物學,2015,34(11):2465-2472.
[36]李 瀅,匡雪君,朱孝軒,等. 長春花密碼子使用偏好性分析[J]. 中國中藥雜志,2016,41(22):4165-4168.
