五加科植物鯊烯合酶核苷酸及其編碼氨基酸序列的生物信息學分析
2019-08-21 01:13:57魏磊常霞全彥濤
江蘇農業科學 2019年12期
魏磊 常霞 全彥濤


摘要:鯊烯合酶(squalene synthase,EC2.5.1.21,簡稱SQS)是三萜類化合物生物合成通路的關鍵酶之一。采用生物信息學方法對13種五加科植物的SQS核苷酸及其編碼氨基酸序列的結構、理化性質、磷酸化位點、親/疏水性、信號肽、導肽、跨膜結構域、亞細胞定位、二級結構、功能域、三級結構及進化關系進行初步預測和分析,并構建SQS蛋白家族的系統進化樹。結果表明,13種五加科植物的SQS氨基酸序列結構與理化性質基本一致,均表現出親水性,沒有信號肽,具有跨膜結構域;亞細胞定位分析顯示,除Panax sokpayensis定位于內質網膜上,其余均定位于質膜上。α螺旋和無規則卷曲為SQS二級結構中最主要的結構元件,保守區包括底物結合區、鎂離子結合位點、活性位點殘基蓋、催化殘基和2個天冬氨酸富集區,具有典型的多聚異戊二烯基合成酶活性結構域和鯊烯/八氫番茄紅素合成酶活性結構域。分析結果可為SQS的酶學特性及三萜類化合物生物合成的分子機制研究提供理論基礎。
關鍵詞:五加科;生物信息學;鯊烯合酶;三萜生物合成
中圖分類號:S188?? 文獻標志碼: A? 文章編號:1002-1302(2019)12-0057-06
三萜類化合物由6個異戊二烯結構單元組成,以共同前體鯊烯(squalene)為底物,在不同三萜合酶的催化下形成的C30類萜類化合物,廣泛分布于植物、動物和微生物中。在植物界,三萜類化合物在五加科、豆科、大戟科、衛矛科等科屬中廣泛存在,且具有抗癌、解熱、抗炎、鎮痛、溶血、抗菌、抗病毒等重要的藥理活性[1]。在植物三萜合成通路(圖1)中,鯊烯合酶(squalene synthase,簡稱SQS)處于法尼基焦磷酸(FPP)到其他產物(二萜、四萜等)的分支點上,催化2分子FPP縮合生成1分子鯊烯,而鯊烯是三萜、甾醇等萜烯類物質生物合成的共同前體,因此,SQS是三萜類化合物生物合成通路中的關鍵酶[2]。
至今,多種植物的SQS已經或正在被深入研究,包括銀杏(Ginkgo biloba,GeneBank:APB03419.1)、紫杉(Taxus cuspidata,GenBank:ABI14439.1)等裸子植物及以煙草(Nicotiana tabacum,GeneBank:AAB08578.1)、擬南芥(Arabidopsis thaliana,GeneBank:AAD00296.1)為代表的被子植物。Floressánchez等用果膠處理貓掌樹懸浮細胞,使得SQS活性提高了3倍以上,進而增加了萜烯類物質產量[3];Han等通過誘導睡茄鯊烯合酶的表達,能夠提高植物甾醇的含量[4];而張毅等反義抑制煙草SQS,降低了植物甾醇的生物合成[5];Patel等抑制人參鯊烯合酶的表達,可使三萜皂苷的生成量降低[6]。以上研究均表明,對植物SQS進行調控可改變后續產物的產量。
五加科(Araliaceae)植物多為古老而又名貴的藥用植物,通常其體內三萜類化合物含量豐富,活性顯著,廣泛應用于醫藥和食品行業,如具有大補元氣、固脫、生津、安神和益智功效的人參(Panax ginseng)[7],活血止血、消腫定痛、金刀跌傷的三七(P. notoginseng)[8],補氣安神、強精滋腎、祛風除濕、活血止痛的楤木(Aralia elata)[9]等。目前,已從五加科人參、三七、楤木、刺五加(Eleutherococcus senticosus)、西洋參(P. quinquefolius)等中藥材中克隆到了SQS的基因。鯊烯合酶作為植物三萜類化合物生物合成過程中的一個關鍵酶,受到越來越多的關注,本研究利用生物信息學方法,篩選出五加科植物中已被克隆的13個SQS蛋白,對其核苷酸及編碼的氨基酸序列的組成、理化性質、磷酸化位點、親/疏水性、信號肽、導肽、跨膜結構域、亞細胞定位、二級結構、結構域、三級結構和進化關系進行預測和分析,以期為更多物種SQS蛋白結構和功能的深入研究提供理論依據。
1 材料與方法
1.1 材料
從美國國立生物技術信息中心(National Center of Biotechnology Information,簡稱NCBI)下載并篩選出了五加科植物人參、三七、西洋參、刺五加、竹節參(Panax japonicus)、楤木和P. sokpayensis的13種完整的SQS核苷酸和氨基酸序列(表1)。
1.2 方法
2 結果與分析
2.1 五加科植物SQS核苷酸及其編碼氨基酸序列的結構和理化性質
利用ORF Finder和ProtParam在線網站對13種五加科植物SQS核苷酸及其編碼氨基酸序列進行理化性質分析。由表2可知,SQS核苷酸序列的起始密碼子均為ATG,終止密碼子均為TGA;氨基酸殘基數(amino acids,aa)在414~446個之間;各蛋白相對分子質量在46 923.26~51 063.97 ku 之間,中位值為47 067.53 ku;理論等電點范圍為5.96~6.67,平均為6.28,說明SQS為酸性蛋白。從氨基酸組成可以看出,13種SQS蛋白所含酸性氨基酸殘基比例均高于堿性氨基酸,進一步說明SQS為酸性蛋白。各種植物SQS蛋白中,含量最豐富的氨基酸殘基主要集中在亮氨酸(Leu)、丙氨酸(Ala)、絲氨酸(Ser)、賴氨酸(Lys)和異亮氨酸(Ile)上。除P. sokpayensis外,其余12種五加科植物總原子數和消光系數基本一致。人參2、人參3、西洋參1、西洋參3、刺五加1、刺五加2不穩定系數低于40,為穩定蛋白,其余均為不穩定蛋白。
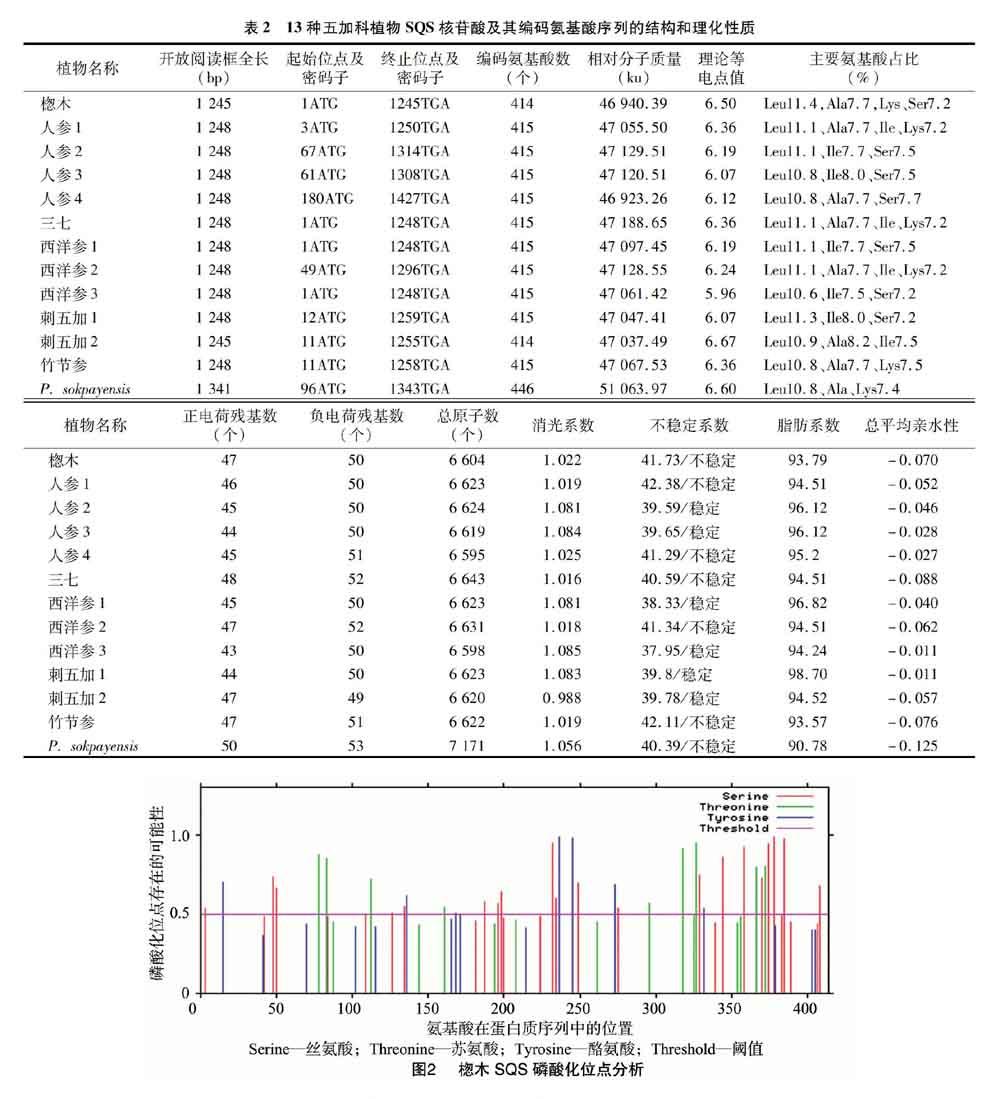
2.2 五加科植物SQS蛋白的磷酸化位點
利用NetPhos 3.1 Server對13種五加科植物SQS蛋白磷酸化位點進行預測,磷酸化位點在17~20個之間。以楤木SQS蛋白為例,超過閾值(Threshold)0.5判定為磷酸化位點,共有36個磷酸化位點(圖2),其中絲氨酸磷酸化位點20個,分別為S3、S48、S50、S126、S134、S187、S196、S198、S232、S234、S249、S215、S329、S344、S358、S370、S374、S378、S385、S408;蘇氨酸磷酸化位點9個,分別為T78、T83、T112、T161、T296、T318、T327、T366、T372;酪氨酸磷酸化位點7個,分別為Y15、Y136、Y168、Y236、Y245、Y273、Y332。除個別位點以外,其余植物SQS蛋白磷酸化位點與楤木基本一致。
2.3 五加科植物SQS的親/疏水性、信號肽、導肽、跨膜結構域和亞細胞定位特征
2.3.1 蛋白親/疏水性的預測 蛋白質親/疏水性氨基酸組成是蛋白質折疊的主要驅動力[10],利用Protscale在線預測親疏水性,以楤木SQS為例,其多肽鏈中第351位氨基酸有最低的親水性分值-2.433,位于395和398位的氨基酸有最高的疏水性分值3.567。其中,親、疏水性氨基酸分別占55.4%和 44.6%,推測折疊的蛋白為親水性蛋白。此外,肽鏈兩端為親水性氨基酸,中間有2段疏水性氨基酸,這與跨膜結構的預測結果相吻合。其余12種SQS的親/疏水性分析結果都與楤木相似。
2.3.2 信號肽的預測和分析 信號肽是新合成的多肽鏈中用于指導蛋白質跨膜轉移(定位)的N-末端的氨基酸序列,一般由16~26個氨基酸組成,在信號肽的引導下,新合成的蛋白質進入內質網腔,而信號肽自身的序列在信號肽酶的作用下被切除[11]。利用在線工具Signal P 4.1 Server的神經網絡算法對13種五加科植物的SQS蛋白進行信號肽的預測,結果表明均不存在信號肽。
2.3.3 導肽的預測和分析 導肽又稱轉運肽,大約18~20個氨基酸長度,位于新合成蛋白質的N端,其含有豐富的堿性氨基酸,這些氨基酸對于蛋白質的定位具有重要的作用[12],能將肽鏈準確地定位于細胞的某一個細胞器或組織上[13]。因此,對導肽的預測分析有助于蛋白質的功能和作用機制研究。通過在線預測工具TargetP 1.1 Server,對五加科植物SQS序列進行預測。以楤木SQS蛋白為例,預測可靠性為Ⅲ級,其葉綠體轉運肽、線粒體靶位肽、分泌途徑信號肽及其他的分值分別為0.065、0.237、0.061、0.743,并且不存在導肽分裂位點。因此,無法確定楤木SQS是否具有導肽。其他11種五加科植物的SQS預測分析結果與楤木完全一致。但P. sokpayensis的SQS蛋白預測可靠性為Ⅳ級,可能含有較低相似度的分泌途徑信號肽,但不存在導肽分裂位點。
2.3.4 跨膜結構域特征 跨膜結構域一般由20個左右的疏水性氨基酸殘基組成,主要形成α螺旋,常由跨膜蛋白的效應區域展現[14]。利用在線工具TMHMM Server v.2.0對13種五加科植物SQS蛋白跨膜結構進行預測可知,SQS蛋白均具有跨膜結構域,P. sokpayensis有1個,其余五加科植物均有2個。跨膜區域位于C端的疏水序列,用于錨定在生物膜上。
2.3.5 亞細胞定位特征 蛋白質在細胞中合成后被轉運到特定的細胞器發揮作用,蛋白質亞細胞定位分析及預測能極大地加速蛋白質結構和功能的研究[15]。采用PSORT Prediction在線工具對13種五加科植物的SQS進行亞細胞定位。由表3可知,P. sokpayensis SQS位于內質網膜上的可能性為0.850,其余植物SQS位于質膜上的可能性均大于0.640。因此判斷SQS P. sokpayensis定位于內質網膜上,其余植物SQS定位于質膜上。
2.4 五加科植物SQS二級結構預測
蛋白質二級結構是指借助于氫鍵沿一維方向排列成具有周期性結構的構象,是多肽鏈局部的空間結構(構象),它們是構成蛋白質高級結構的基本要素。利用SOPMA對13種五加科植物SQS二級結構進行預測。結果顯示,五加科SQS中均有α螺旋、無規則卷曲、延伸鏈和β折疊。以楤木SQS為例,由表4可知,其主要結構元件是α螺旋(68.36%)、β折疊(5.31%)、無規則卷曲(20.05%)和延伸鏈(6.28%)。其余蛋白二級結構的主要結構元件與比例與楤木SQS基本一致。
2.5 蛋白質功能結構域的預測和分析
結構域是蛋白質分子中介于二級與三級結構之間的一種結構層次,是指蛋白質亞基結構中明顯分開的緊密球狀結構區域,是蛋白質三級結構內的獨立折疊單元,具有特定的生物學功能,結構域通常也是蛋白質的功能域[16]。采用NCBI在線軟件Conserved domains對楤木SQS蛋白的氨基酸序列進行保守區和功能域的預測分析,結果表明,楤木SQS蛋白保守區域包括底物結合區、鎂離子結合位點、活性位點殘基蓋、催化殘基和2個天冬氨酸富集區,具有典型的多聚異戊二烯基合成酶活性結構域和鯊烯/八氫番茄紅素合成酶活性結構域,屬于Isoprenoid-Biosyn-C1超家族(圖3)。其余SQS蛋白均存在上述結構域,為類異戊二烯生物合成酶。
x2.6 五加科植物SQS蛋白三級結構的預測和分析
蛋白質的三級結構是在二級結構的基礎上進一步盤繞、折疊形成的天然構象,主要依靠氨基酸側鏈之間的疏水相互作用、氫鍵、二硫鍵、范德華力和靜電作用維持。蛋白質的功能與其三級結構密切相關,對蛋白質三級結構的預測和分析,有助于理解蛋白質結構與功能之間的相關性[17]。利用同源建模工具 SWISS-MODEL對楤木SQS蛋白進行三級結構預
測和分析,找到模板蛋白(ACCESSION:3wef.2.A;Sequence Identity:47.73%;GMQE:0.63),再用Swiss Pdb-Viewer工具顯示楤木SQS結構域的3D結構。結果顯示,楤木SQS為單體蛋白結構,活性中心主要由幾個α螺旋圍繞形成1個穴狀活性中心結構,其20個磷酸化位點位于蛋白質表面。主要結構單元為螺旋結構,伴有折疊、無規則卷曲和延伸鏈結構(圖4),與二級結構預測結果一致。
2.7 SQS蛋白的系統進化樹分析
來源于同一祖先的不同植物在進化過程中的關系可以通過進化樹來描述,通過構建植物進化樹,可以了解一種植物在進化過程中的地位[18]。用MEGA 7.0軟件對包括五加科在內的28種有代表性的SQS蛋白構建系統進化樹。結果顯示,來源于裸子植物、被子植物、動物、細菌、真菌的SQS按照不同類群分為5群,在進化遺傳學上親緣越近的物種,在SQS的分子系統進化樹上距離越近(圖5)。基于氨基酸序列重建的系統進化樹,雖不能完全客觀地反映出植物系統分類學關系和漫長歷史長河中的自然演化進程,但其結果對判斷不同植物之間的親緣關系仍具有一定的借鑒意義[19]。
3 結論與討論
植物萜類化合物的生物合成途徑包括2種:一種是甲戊二羥酸(MVA)途徑,也稱細胞質途徑,主要合成倍半萜、三萜和甾醇等化合物;一種是脫氧木酮糖磷酸酯(DXP)途經,又稱質體途徑,主要合成單萜、二萜、類胡蘿卜素等化合物[20-21]。雖然2種途徑在細胞的不同區域獨立進行,但許多證據表明2種途徑有著密切的聯系,細胞質和質體之間存在某些中間產物交換現象。由于SQS競爭利用FPP來合成三萜及甾醇,會影響單萜、二萜類物質的合成[22],因此SQS含量和活性決定了相關物質的產量。
蛋白一級結構預測分析結果表明,SQS蛋白為酸性的親水性蛋白,部分不穩定,部分穩定,具有明顯的疏水區和親水區,不具有信號肽,可推測SQS不是分泌性蛋白,這與其屬于細胞質中的MVA途徑關鍵酶相一致;都具有2個跨膜結構域,無法確定是否具有導肽,也未發現導肽的分裂位點;亞細胞定位預測分析表明其可能定位在質膜、內質網膜或高爾基體上。SQS蛋白的二級結構均以α螺旋、β折疊、無規則卷曲和延伸鏈分布于整個肽鏈結構中。所有五加科SQS蛋白氨基酸序列中都含有多聚異戊二烯基合成酶的活性結構域和底物結合位點結構域,屬于Isoprenoid-Biosyn-C1超家族,為類異戊二烯生物合成酶。
利用生物信息學的方法對五加科植物SQS氨基酸序列的生理生化特性進行預測和分析,可以為SQS蛋白及其編碼基因的克隆提供可靠的依據;對其序列結構的預測和分析,可為其蛋白表達與修飾提供指導;并為更多物種SQS蛋白及其編碼基因的克隆提供可靠的依據。對其二級及高級結構的預測和分析有利于深入探討該酶結構和功能之間的關系、作用機制和代謝過程。
參考文獻:
[1]劉江亭,蔣海強. 三萜類化合物藥理作用研究進展[J]. 中國中醫藥咨訊,2011,3(10):321.
[2]Goldstein J L,Brown M S. Regulation of the mevalonate pathway[J]. Nature,1990,343(6257):425.
[3]Floressánchez I J,Ortegalópez J,Ramosvaldivia A C. Biosynthesis of
sterols and triterpenes in cell suspension cultures of Uncaria tomentosa[J]. Plant & Cell Physiology,2002,43(12):1502.
[4]Han J Y,In J G,Kwon Y S,et al. Regulation of ginsenoside and phytosterol biosynthesis by RNA interferences of squalene epoxidase gene in Panax ginseng[J]. Phytochemistry,2010,71(1):36-46.
[5]張 毅,劉 彥,王 紅,等. 轉青蒿反義鯊烯合酶基因對煙草鯊烯合酶基因表達的影響[J]. 農業生物技術學報,2005,13(4):416-422.
[6]Patel N,Patel P,Kendurkar S V,et al. Overexpression of squalene synthase in Withania somnifera,leads to enhanced withanolide biosynthesis[J]. Plant Cell Tissue & Organ Culture,2015,122(2):409-420.
[7]王 月,翟華強,魯利娜,等. 人參的本草考證及現代研究綜述[J]. 世界中醫藥,2017,12(2):470-473.
[8]孫千惠,劉海嬌,楊小玉,等. 三七本草考證[J]. 中醫藥信息,2017,34(5):113-117.
[9]趙俊男,譚玉婷,姜天童,等. 龍牙楤木藥理作用研究進展[J]. 吉林中醫藥,2016(2):207-210.
[10]張 弢. 不同植物查爾酮合成酶CHS基因的生物信息學分析[J]. 江西農業學報,2012,24(6):5-8.
[11]翟中和,王喜忠,丁明孝. 細胞生物學[M]. 北京:高等教育出版社,2000:191.
[12]強 毅. 植物蔗糖磷酸合成酶的生物信息學分析[J]. 現代生物醫學進展,2007,7(4):557-560,570.
[13]金元昌,向育君,李景鵬. 重組人促性腺激素釋放激素及導肽的分離純化與活性分析[J]. 生物技術通報,2008(3):87-90,94.
[14]董 嬌,周 軍,辛培堯,等. 不同植物LDOX/ANS基因的生物信息學分析[J]. 基因組學與應用生物學,2010,29(5):815-822.
[15]Herr D. Secretion of cellulase and β-glucosidase by Trichoderma viride ITCC-1433 in submerged culture on different substrates[J]. Biotechnology & Bioengineering,2010,21(8):1361-1371.
[16]王鏡巖,朱圣庚,徐長法. 生物化學[M]. 北京:高等教育出版社,2002:533.
[17]蔡娜娜,陳月輝,李 偉. 基于神經網絡的蛋白質三級結構預測[J]. 計算機工程,2010,36(9):176-177.
[18]于 劍,葉 齊,寧書菊,等. 馬藍等79種植物分支酸合成酶的生物信息學分析[J]. 中國中藥雜志,2018,43(4):721-730.
[19]吳春太,李維國,高新生,等. 植物DNA甲基轉移酶的生物信息學分析[J]. 西南大學學報(自然科學版),2010,32(4):83-89.
[20]Bouvier F,Rahier A,Camara B. Biogenesis,molecular regulation and function of plant isoprenoids[J]. Progress in Lipid Research,2005,44(6):357-429.
[21]Liao P,Hemmerlin A,Bach T J,et al. The potential of the mevalonate pathway for enhanced isoprenoid production[J]. Biotechnology Advances,2016,34(5):697-713.
[22]Shechter I. The road to squalene synthase[J]. Biochemical & Biophysical Research Communications,2002,292(5):1261-1266.楊旭穎,安麗君. 擬南芥細胞周期調控因子KRP2的原核表達及蛋白純化[J]. 江蘇農業科學,2019,47(12):63-65.
