基于ISSR標記的鐵線蓮園藝品種遺傳多樣性分析和指紋圖譜構建
2019-08-21 01:13:57和文志潘鵬舉楊玲
江蘇農業科學 2019年12期
和文志 潘鵬舉 楊玲


摘要:為進一步明確鐵線蓮屬16個園藝品種之間的親緣關系和遺傳多樣性,利用12條ISSR引物對16份材料進行PCR擴增,共獲得117條清晰的條帶,多態性條帶占比為97.44%。利用引物UBC815和UBC817構建16個鐵線蓮品種的指紋圖譜,可以有效地對鐵線蓮進行區分和鑒定。利用NTSYS軟件計算出鐵線蓮品種間的遺傳相似系數為 0.20~0.73,平均相似系數為0.51。通過UPGMA法進行聚類分析,在相似系數為0.53時,可將16份材料劃分為4大類。結果表明,鐵線蓮品種之間的遺傳關系較為復雜,利用ISSR分子標記技術可以從分子水平揭示16份材料的遺傳多樣性水平和親緣關系。
關鍵詞:鐵線蓮;品種;ISSR;遺傳多樣性;指紋圖譜
中圖分類號: Q756;S682.390.3? 文獻標志碼: A? 文章編號:1002-1302(2019)12-0079-04
鐵線蓮為毛茛科(Ranunculaceae)鐵線蓮屬(Clematis L.)植物,現已知的鐵線蓮約有300種,廣泛分布于世界各地。我國約有133種,主要分布在華中和西南地區[1]。鐵線蓮具有非常高的藥用價值和觀賞價值。該屬植物不僅花型多樣,有平盤形、十字形、鐘形、長瓣形等,還兼具花期長、色彩豐富的特點,擁有“藤本皇后”的美譽。在園藝家和植物學家幾百年的栽培和育種過程中,培育出了上千個各具特色的鐵線蓮園藝品種[2]。這些品種中,絕大部分都是由多個不同的種雜交而來,還有部分品種是由野生種馴化而來[3]。長時間的人工選擇過程中,鐵線蓮品種的遺傳背景和各品種之間的親緣關系變得模糊不清,有些雜交品種已經找不到最初的雜交親本,這給雜交育種親本選擇帶來一定的困難。同時,鐵線蓮很多品種外觀形態相似度高,導致鐵線蓮品種鑒定越來越難。由于受環境影響因素較大,傳統形態學分類方法難以對鐵線蓮品種進行有效區分。因此,運用分子生物學手段對鐵線蓮品種遺傳多樣性和親緣關系進行研究,可為鐵線蓮品種在分子水平的分類與鑒定提供依據。目前,在有關鐵線蓮品種遺傳多樣性研究上,前人多從形態學和細胞學[4-7]等方面開展了一些研究,利用分子生物學手段進行相關研究的報道較少。
簡單重復間序列(inter-simple sequence repeat,ISSR)標記技術具有操作簡單、穩定性好和多態性豐富等特點,廣泛應用于植物品種鑒定、基因作圖與定位、遺傳多樣性和系統發育等的研究。孫正海等建立了鐵線蓮ISSR-PCR的反應體系,并且進行反應體系優化和引物篩選,但未進一步研究該分子標記技術在鐵線蓮品種遺傳多樣性研究中的可行性與穩定性[8-9]。本研究利用ISSR分子標記技術從分子水平對鐵線蓮16個園藝品種遺傳多樣性進行探究,為ISSR分子標記技術在鐵線蓮品種分類鑒定、親緣關系分析以及種質資源利用和新品種培育過程中親本選擇提供理論依據。
1 材料與方法
1.1 材料
試驗材料為2015年12月從國外引進的16個鐵線蓮園藝品種。試驗地為西南林業大學一號教學樓前苗圃,位于云南省昆明市盤龍區,屬于北亞熱帶低緯高原山地季風氣候,年平均氣溫16.5 ℃,年均降水量1 450 mm,無霜期278 d。采集生長良好的植物幼嫩葉片,置于放入硅膠的自封袋當中,帶回實驗室置于-80 ℃冰箱保存備用。品種信息(來自世界園藝鐵線蓮協會)詳見表1。
1.2 方法
1.2.1 DNA提取與檢測 采用DNA提取試劑盒(OMEGA D2485-01)提取植物基因組DNA。利用Nanodrop ND-2000微量核酸蛋白檢測儀檢測DNA質量及濃度。利用0.8%瓊脂糖凝膠電泳檢測DNA鏈的完整性,電泳緩沖液為 0.5×TBE。
1.2.2 ISSR引物篩選與PCR擴增 試驗所選100條引物(UBC801~UBC900)由哥倫比亞大學公布,昆明碩擎生物科技有限公司合成。PCR反應體系為25 μL,包括模板DNA,引物2 μL,12.5 μL PCR反應混合物(Master Mix),加入雙蒸水使體系達到25 μL。反應程序:94 ℃預變性3 min;94 ℃變性90 s, 50 ℃退火30 s, 72 ℃延伸1 min,43個循環;72 ℃延伸
從上述100條引物中初步篩選出28條進行退火溫度梯度的試驗,篩選出條帶數量豐富、清晰穩定的引物12條,并確定最適退火溫度。最后,利用這12條引物對16個鐵線蓮品種進行ISSR-PCR擴增,使用凝膠成像系統進行拍照分析。
1.2.3 數據記錄與分析 根據凝膠成像結果,同一引物擴增出來的條帶,在同一遷移率上出現條帶記為“1”,丟失條帶記為“0”,建立0、1二元數據矩陣。選擇多態性位點比較豐富的引物,構建16個鐵線蓮品種的指紋圖譜;利用NTSys2.1數據分析軟件計算品種間的遺傳距離和遺傳相似系數,并采用UPGMA法構建其遺傳聚類圖。
2 結果與分析
2.1 鐵線蓮DNA提取效果
利用DNA提取試劑盒提取鐵線蓮16個品種的DNA都是一條完整清晰的譜帶(圖1),沒有RNA污染、拖尾和彌散現象,利用超微量核酸蛋白測定儀測得DNA的濃度為55~120 ng/μL。提取的DNA可以滿足ISSR-PCR反應對模板DNA的質量要求。
2.2 ISSR引物篩選及擴增結果
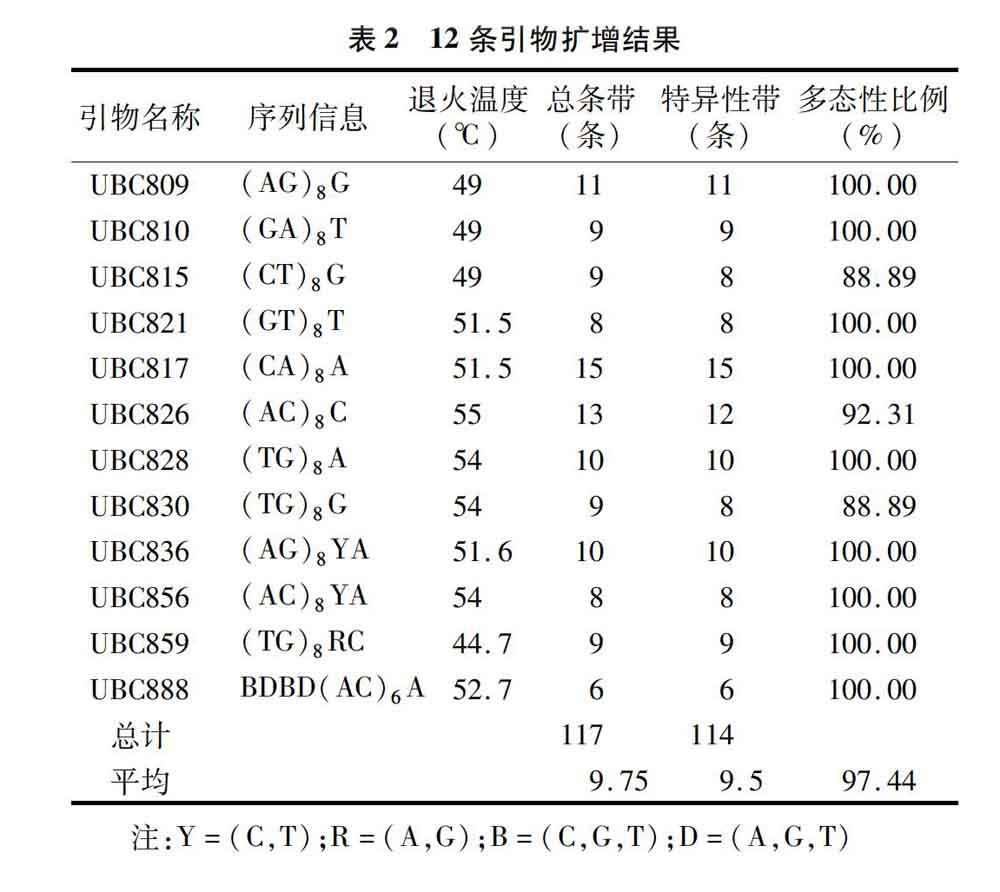
從100條ISSR引物中篩選出12條擴增條帶清晰穩定、數量豐富的引物,分別為UBC809、UBC810、UBC815、UBC821、UBC817、UBC826、UBC828、UBC830、UBC836、UBC856、UBC859、UBC888。通過退火溫度梯度(44.7~62.5 ℃)試驗,確定了每條引物的最佳退火溫度。利用篩選出的引物對16個供試材料進行ISSR-PCR擴增。12條引物共擴增出117條帶,其中特異性條帶共有114條,每個引物平均擴增出來9.75個位點,平均特異性位點達到9.5個,多態性比率達到97.44%(表2);擴增出的條帶片段大小為250~1 500 bp。引物UBC817、UBC815和UBC856擴增結果見圖2、圖3、圖4。
x2.3 16個鐵線蓮園藝品種的ISSR聚類結果
利用12條引物擴增出來的117譜帶建立16個鐵線蓮品
種的遺傳相似系數矩陣(表3),各品種之間的相似系數在 0.20~0.73之間,平均相似系數為0.51。說明鐵線蓮品種具有較高的遺傳多樣性,各品種間的遺傳變異較大,環境適應能力較強。其中,包查德女伯爵和紅衣主教相似系數最大,為0.73,這表明2個品種之間的親緣關系相對較近;塞西爾和邁克萊特相似系數最小,為0.20,表明這2個品種的親緣關系相對較遠。從遺傳相似系數來看,塞西爾同其他品種之間的遺傳相似系數較小,為0.20~0.35,表明該品種與其他品種相比遺傳變異大、親緣關系遠。
采用UPGMA法對供試的16個鐵線蓮園藝品種進行聚類分析,結果表明,在相似系數為0.50處,可將16份材料聚為3類(圖5)。其中,塞西爾劃為A類,紫鈴鐺劃為B類,C類包括其余14個品種。在相似系數為0.53處,可將16份材料劃分為4個類群。塞西爾劃為第1類;紫鈴鐺劃為第2類;第3類包括月光、邁克萊特、格拉芙泰美女和普魯吐斯;其余10個品種劃為第4類。
2.4 16個鐵線蓮園藝品種的ISSR指紋圖譜構建
利用擴增出條帶清晰、條帶數量多、多態性豐富的引物構建16個鐵線蓮品種的遺傳指紋圖譜。利用引物UBC815和UBC817構建16個鐵線蓮品種的指紋圖譜(表4),該指紋圖譜可以有效地對這些鐵線蓮品種進行區分、鑒定。
3 結論與討論
本研究利用ISSR分子標記技術篩選出的12條引物擴增出117個位點,多態性條帶的比例為97.44%,每條引物擴增出的條帶數為6~15條,多態性條帶所占比例為88.89%~100%。其中12條引物中有9條引物擴增出的條帶多態性達到了100%,說明鐵線蓮栽培品種具有較高的遺傳多樣性和復雜的遺傳背景。利用引物UBC815和UBC817建立起的16個鐵線蓮品種的DNA指紋圖譜可以對不同品種進行有效地區分和快速鑒定。對材料進行ISSR聚類分析,在相似系數0.50處可將16份材料聚為3類。A類只有品種塞西爾;B類只有紫鈴鐺;C類有普魯吐斯、格拉芙泰美女、邁克萊特、月光、美佐世、天真一瞥、小鴨、鉆石球、艾米莉亞花盤、藍天使、包查德女伯爵、紅衣主教、面白和吉恩斯。其中,包查德女伯爵與紅衣主教之間的相似系數最大,為0.73,親緣關系最近,遺傳背景相似。從花朵形態特征來看,包查德女伯爵和紅衣主教除花朵顏色不同外,都具有相同數量苞片,花期都為6—9月,花型都是平盤型,萼片中部具有豎狀條紋,說明ISSR分子聚類與形態聚類在一定程度上的一致性。C類中鐵線蓮的花型都為平盤型,A類花型為長瓣型,B類為鈴鐺型。以此來看,按照花型分類也可以將材料劃分為3類,這與ISSR聚類結果保持一致,擁有相同花型的品種之間可能具有相對較近的親緣關系。但是在同一花型中,不同品種的花朵顏色、單重瓣及萼片數量等性狀表現不一,這可能是鐵線蓮品種之間相互雜交導致種質資源混雜的結果,說明分子聚類結果與形態學分類結果存在一定的差異。塞西爾和邁克萊特之間的相似系數最小,為0.20,說明這2個品種之間的親緣關系較遠。在鐵線蓮新品種培育中,可以利用這2個品種進行雜交,可能會產生較好的雜種優勢。
按照栽培品種的生境特點、開花特性及開花時間等,園藝家將鐵線蓮品種劃為不同的品種類群。按照ISSR分子標記聚類結果將16份材料劃分的3大類群中,A類只有1個長瓣型品種;B類只有1個單葉型品種;C類中包括8個早花大花型品種、1個德克薩斯型品種、2個意大利型品種和3個晚花大花型品種。ISSR聚類結果顯示,不同園藝類型的鐵線蓮品種劃分到同一類中,表明按照園藝性狀分類結果與分子標記聚類結果存在一定的差異。這主要是因為園藝家對鐵線蓮品種的分類是為了園藝種植的考量,分類依據主要是植物生長環境、適應能力、花朵大小、顏色和開放時間等園藝特性。分子聚類與園藝分類的差異體現了鐵線蓮品種之間的遺傳變異程度之大,這是長期人工選擇和自然進化的共同作用的結果。ISSR聚類結果可以為鐵線蓮品種親緣關系鑒定和雜交育種中親本選擇提供參考。與形態學標記相比,以遺傳物質DNA為基礎的ISSR分子標記重復性好,穩定性高,受環境因素影響小,在植物花卉種源信息鑒定、品種分類和遺傳多樣性評價等[10-12]方面具有廣泛的應用。
參考文獻:
[1]中國科學院中國植物志編委會.中國植物志(第28卷)[M]. 北京:科學出版社,2000.
[2]李同水. 藤本皇后鐵線蓮[M]. 長春:吉林科學技術出版社,2012.
[3]Gardner N,Hokanson S C. Intersimple sequence repeat fingerprinting and genetic variation in a collection of Clematis cultivars in commercial germplasm[J]. HortScience,2005,40(7):1982-1987.
[4]王 磊,王紅梅,賈 君,等. 鐵線蓮屬植物品種及野生種的物候期觀察分析[J]. 浙江農業科學,2016,57(2):198-201.
[5]孟 雪,郝笑微,王 錦. 鐵線蓮8個小花品種的適生性比較[J]. 安徽農業科學,2011,39(35):21597-21599.
[6]高露璐,李林芳,馬育珠,等. 鐵線蓮品種群的花期觀賞性狀分析[J]. 園藝學報,2017,44(5):921-932.
[7]任佳倫. 鐵線蓮品種開花生物學特性及核型分析[D]. 杭州:浙江農林大學,2016.
[8]孫正海,王 錦,李世峰,等. 滑葉藤ISSR-PCR反應體系建立及優化[J]. 云南農業大學學報(自然科學版),2012,27(5):746-750.
[9]王 楠,王 錦,李宗艷,等. 鐵線蓮園藝品種ISSR-PCR反應體系優化與引物篩選[J]. 北方園藝,2016(1):80-83.
[10]張俊杰,馬 旭,廖 源,等. 昆明56個桂花品種ISSR分析[J]. 西部林業科學,2015,44(3):45-52.
[11]羅 清,盧業飛,於艷萍,等. 24份杜鵑屬植物的ISSR分子鑒定[J]. 西北林學院學報,2016,31(3):154-158.
[12]苗美美,趙明明,劉緒明,等. 百合(Lilium spp.)雜交后代ISSR分析親緣關系[J]. 分子植物育種,2016(2):437-441.王連喜,王 田,李 琪,等. 基于作物水分虧缺指數的河南省冬小麥干旱時空特征分析[J]. 江蘇農業科學,2019,47(12):83-88.
