大麥單株產量性狀的雜種優勢及其穩定性分析
2019-08-22 09:54:24孫遠東朱昊華張新忠郭寶健王菲菲許如根
麥類作物學報 2019年8期
孫遠東,朱昊華,呂 超,張新忠, 郭寶健,王菲菲,許如根
(1.江蘇省作物遺傳生理重點實驗室/植物功能基因組學教育部重點實驗室/江蘇省作物基因組學和分子育種重點實驗室, 揚州大學農學院,江蘇揚州 225009;2.江蘇省糧食作物現代產業技術協同創新中心,揚州大學,江蘇揚州 225009)
雜種優勢利用是提高作物產量及改良品質的有效手段之一,水稻、玉米等作物雜種優勢的成功利用為世界糧食產量的增加做出了巨大的貢獻。大麥是世界上的第四大禾谷類作物。1941年,Immer[1]首次報道雜交大麥產量優勢為27%左右;而Hagberg[2]則認為,雜交大麥產量無優勢。Fejer等[3]研究指出,春性與冬性大麥雜交后代的雜種優勢大于100%。林佩霞等[4]研究表明,大麥單株粒重、單株粒數、主穗粒重、分蘗數、粒重的雜種優勢為10%~20%。許如根等[5-6]認為,大麥雜種F1普遍存在中親優勢,僅有28%的組合存在顯著超親優勢。陳曉東等[7]研究認為,大麥株高、穗下節間長、穗長和千粒重多為正向優勢,單株穗數、穗粒數和單株粒重多呈負向優勢。張新忠等[8]的研究表明,72個雜交大麥的7個性狀顯著正向中親優勢組合出現率為72.4% ,顯著超親優勢組合出現率為19.6%。以上研究均是以遺傳背景差異較大的品種(系)或大麥不育系與恢復系配制的雜交種F1為材料,因供試材料的遺傳背景及研究環境不同,組合數偏少,不同研究者報道的大麥雜種優勢的表現及其穩定性差異較大。為明確不同環境下大麥產量性狀的雜種優勢穩定性,本研究以113個(Nasonijo×泰興 9425)DH系配制的永久F2群體及其親本為材料,通過3年4種環境的大田鑒定,研究大麥單株產量性狀雜種優勢的表現及其穩定性,以期為大麥單株產量性狀雜種優勢的QTL分析奠定基礎,也為大麥雜種優勢的利用提供理論依據。
1 材料與方法
1.1 試驗材料
以113個(Nasonijo×泰興 9425)DH系按照1×2、1×3、2×3、2×4、……112×1、112×113、113×1、113×2方式組配的226個雜種F1構建的永久F2(IF2)群體及其親本為材料。其中,Nasonijo為日本引進二棱優質啤酒大麥品種,泰興 9425為江蘇育成的二棱大麥品種,(Nasonijo×泰興 9425)DH系由澳大利亞塔斯馬尼亞大學周美學教授提供。
1.2 試驗設計
本試驗2015-2016年大麥生長季同時在揚州大學試驗農場和鹽城大中農場農科所進行,2016-2017年及2017-2018年大麥生長季均在鹽城大中農場農科所進行。3年4種環境條件下的播種方式相同,親本及雜交種均采取點播,每材料點播3行,行長90 cm,行距20 cm,株距5 cm,3次重復。田間管理同大田生產。
1.3 性狀測定
植株成熟后,每材料從中間行取6株進行考種,田間調查主穗粒數(GNPS)和單株穗數(SNPP),收獲曬干后測單株生物量(BWPP),再用單株脫粒機脫粒,測千粒重(TKW)和單株產量(GWPP)。
1.4 數據統計分析
試驗數據用Excel整理。中親優勢值和超親優勢值參照許如根等[5]方法進行計算,MPH=F1-(P1+P2)/2,BPH=F1-Pb,其中MPH和BPH分別代表中親優勢值和超親優勢值,F1代表雜種性狀表現,P1和P2分別代表雙親性狀表現,Pb代表優值親本表現。描述性統計分析、獨立樣本t測驗、方差分析及聚類分析均使用SPSS 16.0軟件進行。
2 結果與分析
2.1 DH群體和永久F2的產量性狀的平均表現與差異
由表1可知, 2017-2018年鹽城市大中農場農科所試點(E4)IF2群體和DH群體的單株穗數、單株產量及單株生物量等被測產量性狀均大于在其他3個環境的表現,其余3個環境間各被測產量性狀的表現差異不大。IF2群體和DH群體的單株穗數、單株產量及單株生物量在4個環境下的變異系數均大于10%,主穗粒數和千粒重在4個環境下的變異系數均小于10%(DH群體E2環境下的千粒重除外)。除E2環境的單株穗數、E1環境的單株產量及E1和E2環境的單株生物量外,其余各環境下IF2被測產量性狀表現均與DH群體差異顯著。
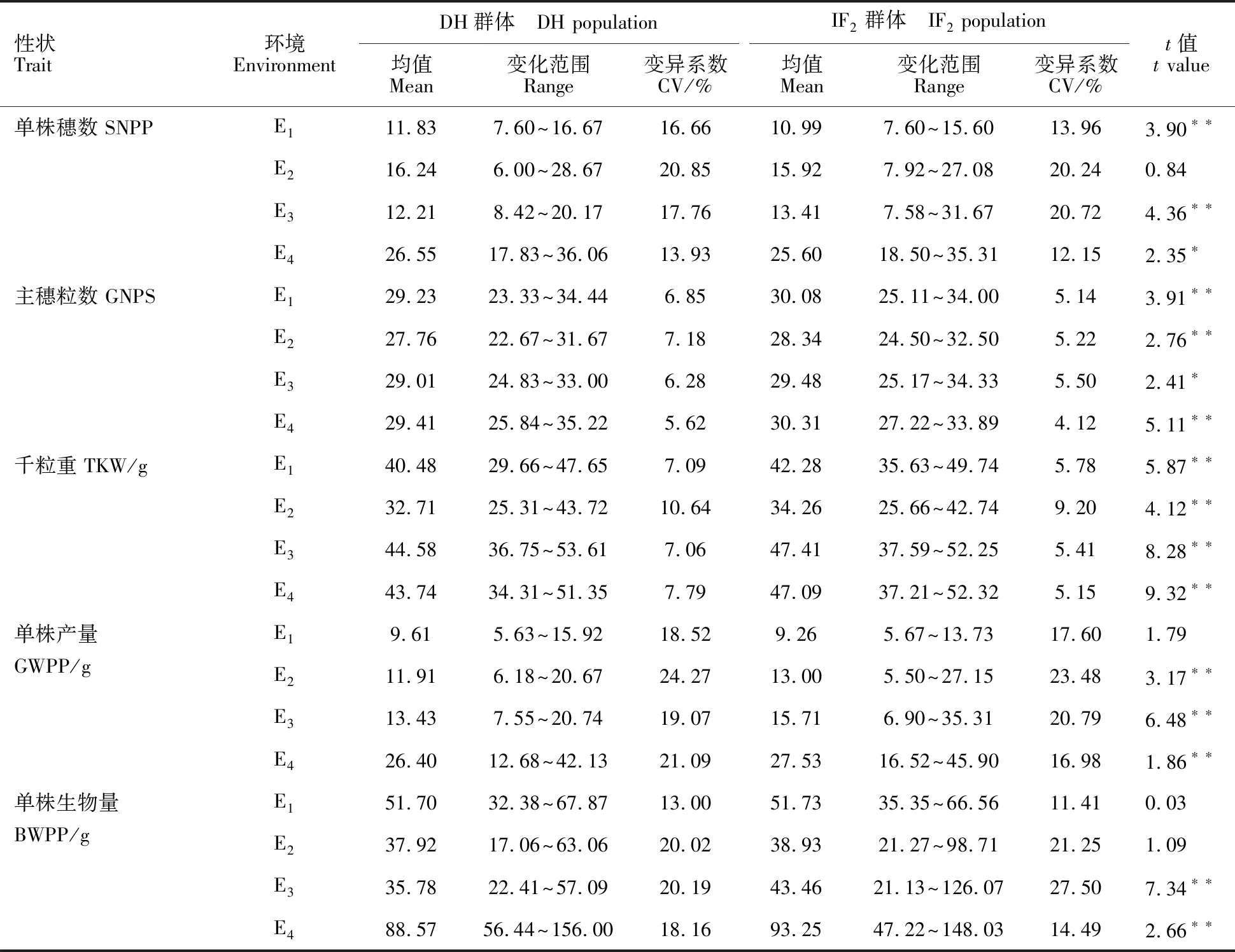
表1 DH群體和永久F2(IF2)群體單株產量性狀的比較Table 1 Comparison of yield traits in DH population and IF2 population
E1:2015-2016年鹽城市大中農場農科所試點; E2:2015-2016年揚州大學試驗農場試點; E3:2016-2017年鹽城市大中農場農科所試點; E4:2017-2018年鹽城市大中農場農科所試點。 *和**分別表示DH群體與其IF2群體在0.05和0.01水平上差異顯著。下同。
E1:2015-2016 at Yancheng city Dazhong Farm; E2:2015-2016 at Yangzhou University; E3:2016-2017 at Yancheng city Dazhong Farm; E4:2017-2018 at Yancheng city Dazhong Farm.*and ** represent significant differences between DH population with their IF2population at 0.05 and 0.01 levels, respectively. The same in tables 2 and 4.
2.2 DH群體和IF2群體產量性狀方差分析
由表2可知,環境、基因型及基因型與環境互作對DH群體和IF2群體的單株穗數、主穗粒數、千粒重、單株產量及單株生物量均有顯著或極顯著影響。說明大麥單株產量性狀表現主要由遺傳因素決定,試點的生態條件和生產條件對其也有顯著影響。
2.3 產量性狀的雜種優勢表現
從表3可知,從雜種優勢的均值和變化范圍來看,E3環境下的單株穗數、單株產量及單株生物量的中親優勢值和超親優勢值、E4環境下的主穗粒數及千粒重的中親優勢值和超親優勢值最大;E4環境下的單株穗數、E2環境下的千粒重、E1環境下的單株產量與單株生物量的中親優勢值和超親優勢值最小,E3和E2環境下的主穗粒數的中親優勢值和超親優勢值最低;說明雜種大麥雜種優勢的表現因試點的生態條件和氣候條件而異。雜種優勢高的試點其正向組合數也較多,低的試點正向組合數則較少。本試驗條件下大麥單株產量被測性狀的雜種優勢在不同環境下的變異系數均較大。
2.4 產量性狀的中親優勢和超親優勢方差分析
由表4可知,環境對單株穗數、千粒重、單株產量和單株生物量的中親優勢和超親優勢有極顯著影響,對主穗粒數的中親優勢和超親優勢影響不顯著;基因型對主穗粒數、千粒重、單數產量和單株生物量的中親優勢和超親優勢有極顯著影響,對單株穗數的中親優勢有顯著影響,而對超親優勢的影響則不顯著;說明雜種大麥單株產量性狀的雜種優勢不僅受組合基因型的影響,還受試點的生態條件和氣候條件影響。
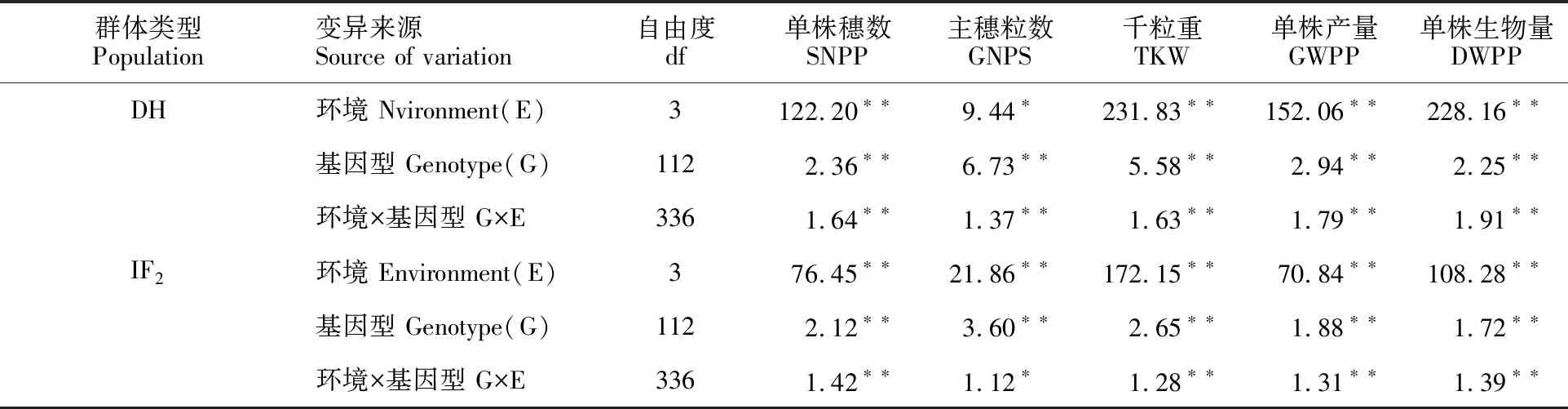
表2 DH群體和IF2群體單株產量性狀的方差分析Table 2 ANOVA of yield traits in DH population and IF2 population
*:P<0.05;**:P<0.01.
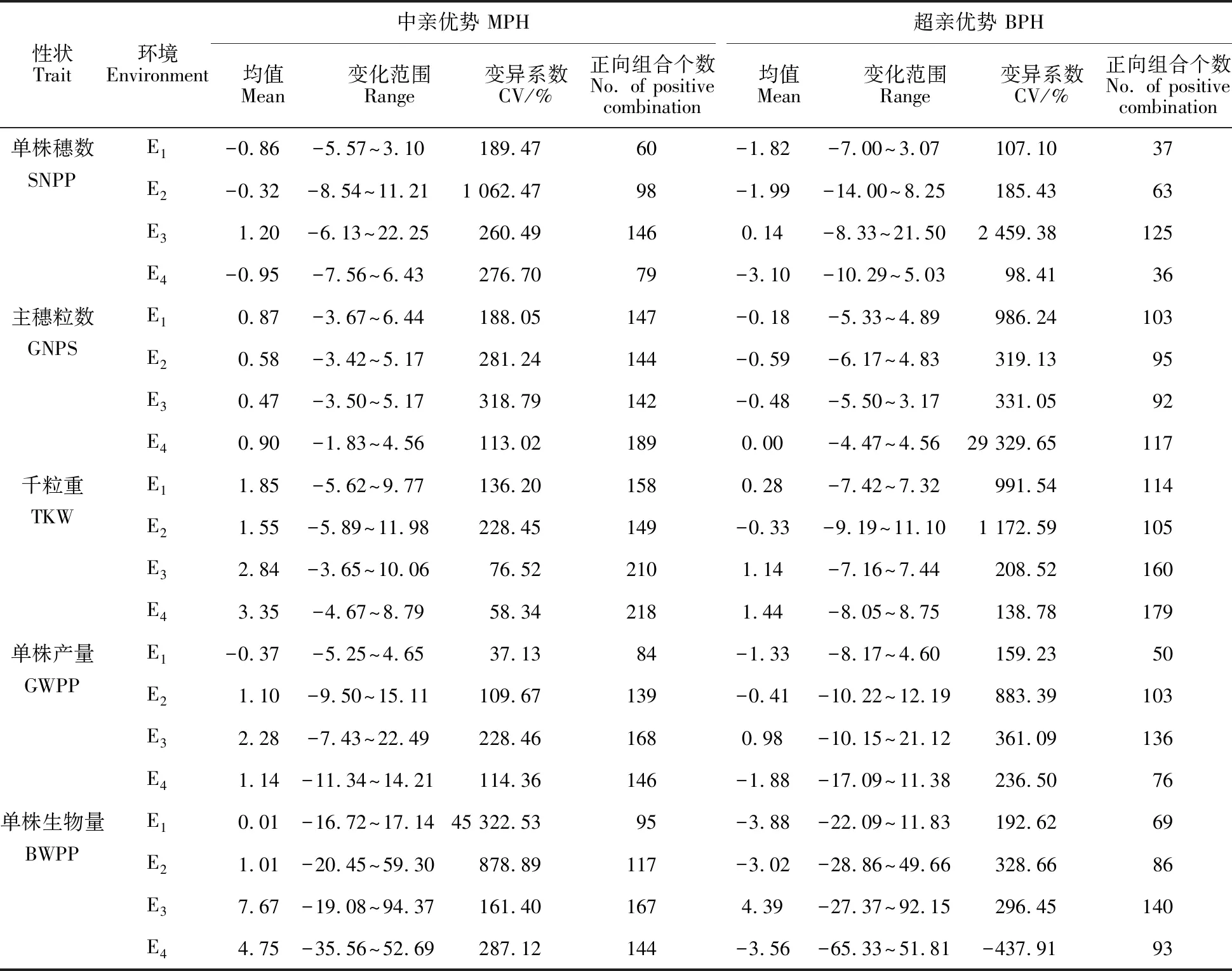
表3 不同環境大麥單株產量性狀的中親優勢與超親優勢表現Table 3 MPH(mid-parents heterosis) and BPH(better-parents heterosis) performance of yield traits in foure nvironments
2.5 產量性狀雜種優勢的穩定性分析
2.5.1 產量性狀雜種優勢穩定性組合數分析
根據不同性狀在4個環境中的雜種優勢值的正負,將其分為4環境正向類、3環境正向類、2環境正向類、1環境正向類和0環境正向類,在不同環境中雜種優勢表現相同的越多,說明其穩定性越高,即4環境和0環境正向類穩定性較高。從表5和表6可以看出,5個性狀在4個環境中穩定性高的組合數較少。中親優勢中,除主穗粒數和千粒重4環境正向類的組合比例較高外,其他3個性狀高穩定性組合比例均小于20%;超親優勢中,穩定性較高組合的比例均小于20%。表明大麥單株產量性狀雜種優勢在環境間的穩定性較差,僅有少數雜交組合穩定性較好。
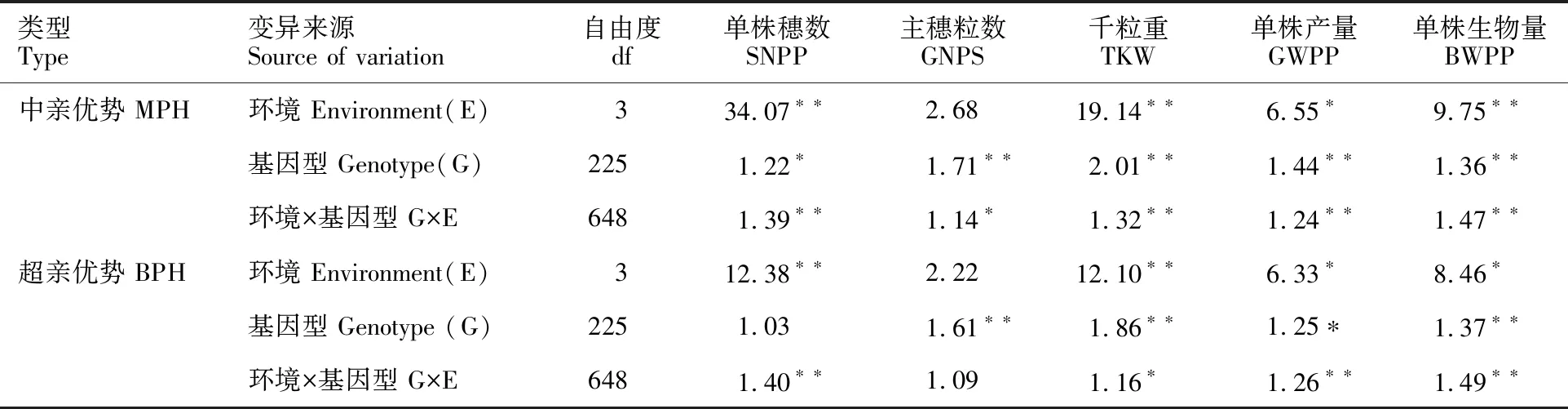
表4 大麥雜種單株產量性狀中親優勢和超親優勢的方差分析Table 4 ANOVA of yield traits for MPH and BPH
*:P<0.05;**:P<0.01.
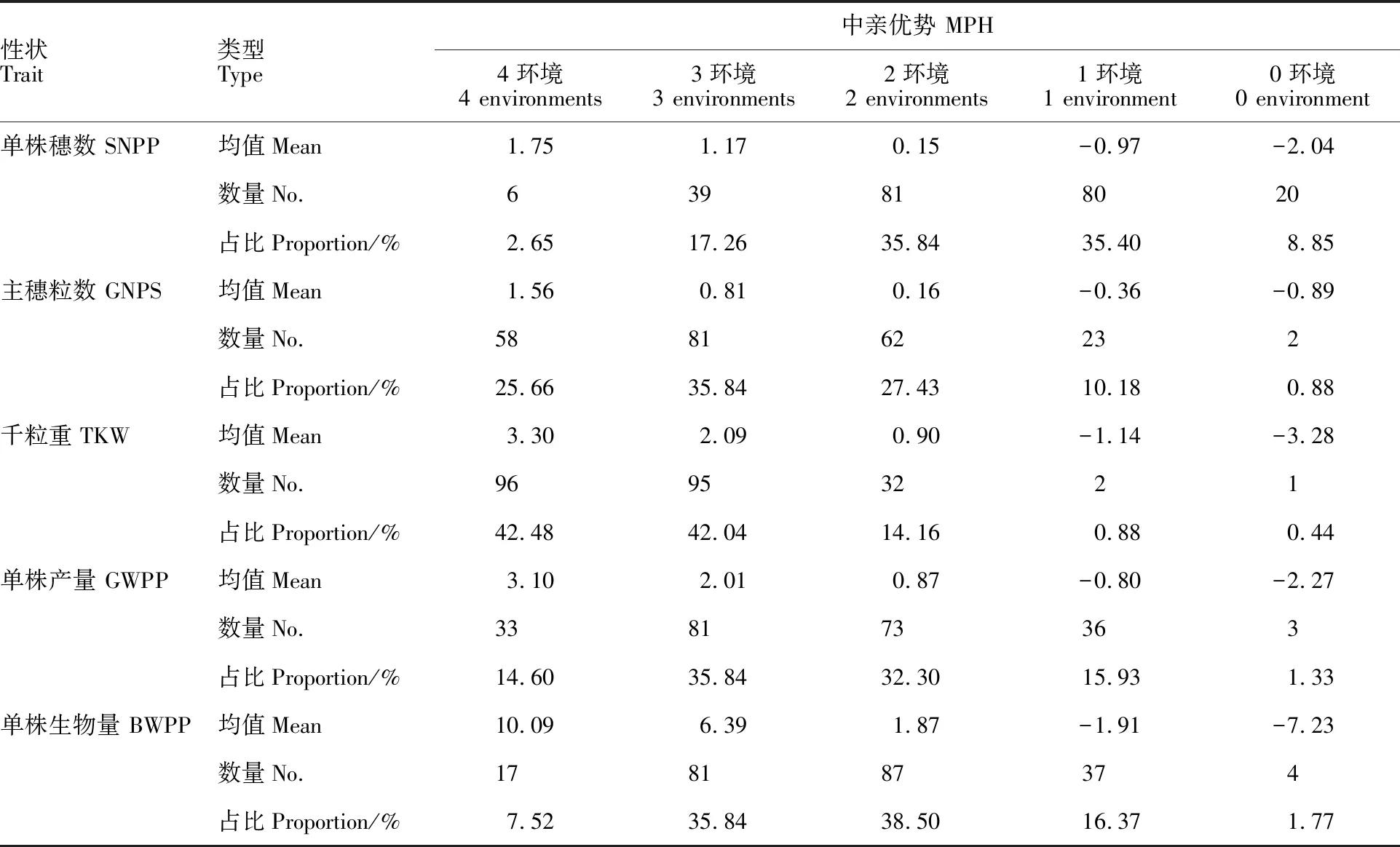
表5 不同環境數大麥單株產量性狀正向中親優勢組合數及中親優勢表現Table 5 No. of positive MPH and performance of MPH in yield traits in four environments
2.5.2 產量性狀中親優勢和超親優勢的強弱 分類
依據大麥單株產量性狀的中親優勢值和超親優勢值的大小,使用SPSS 16.0的K—MEANS聚類法將參試組合分成強、中、弱3類,統計結果如表7。從表7可知,中親優勢中,強類的中親優勢均值變化范圍為1.01~24.41,單株穗數、千粒重和單株產量的中親優勢在E2環境最高,主穗粒數和單株生物量分別在E1和E3環境最高。中類的中親優勢均值變化范圍為-1.13~9.00,除主穗粒數在E1環境中的中親優勢均值最高外,其余4個性狀的中親優勢均值均在E3環境最高。弱類的中親優勢均值變化范圍為-12.39~0.54,單株穗數、單株產量和單株生物量的中親優勢均值在E3環境最高,主穗粒數和千粒重的中親優勢均值在E4環境最高。超親優勢中,強類的超親優勢均值均為正向,單株穗數和單株產量在E3環境、主穗粒數在E1環境及千粒重和單株生物量在E2環境的超親優勢均值最高;中類的超親優勢組合和弱類的超親優勢均值在各環境多為負值。說明雜種大麥普遍存在中親優勢,超親優勢組合相對較少,且雜種優勢在不同環境表現不盡相同。
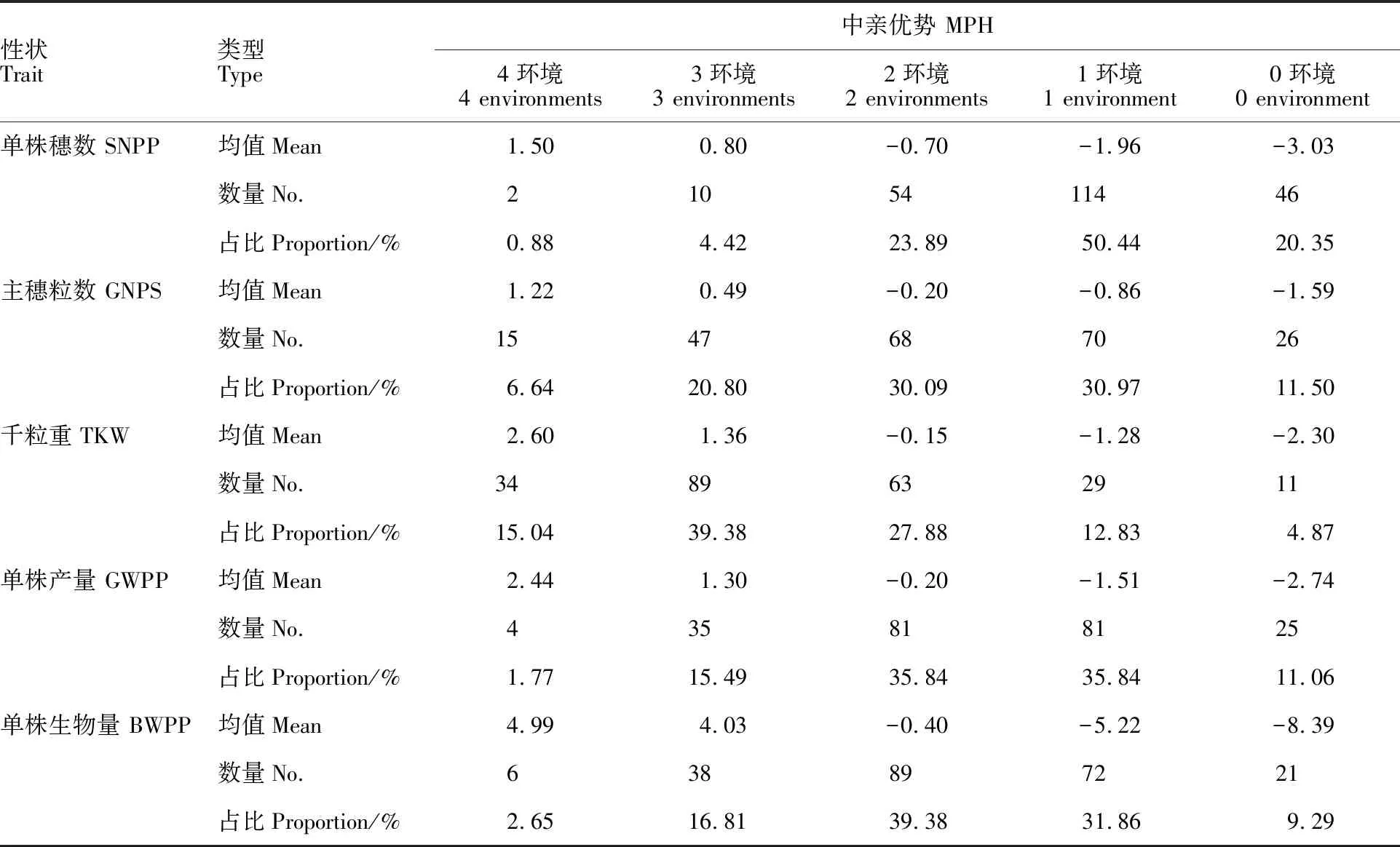
表6 不同環境數大麥單株產量性狀正向超親優勢組合數及超親優勢表現Table 6 No. of positive BPH and performance of BPH in yield traits in four environments
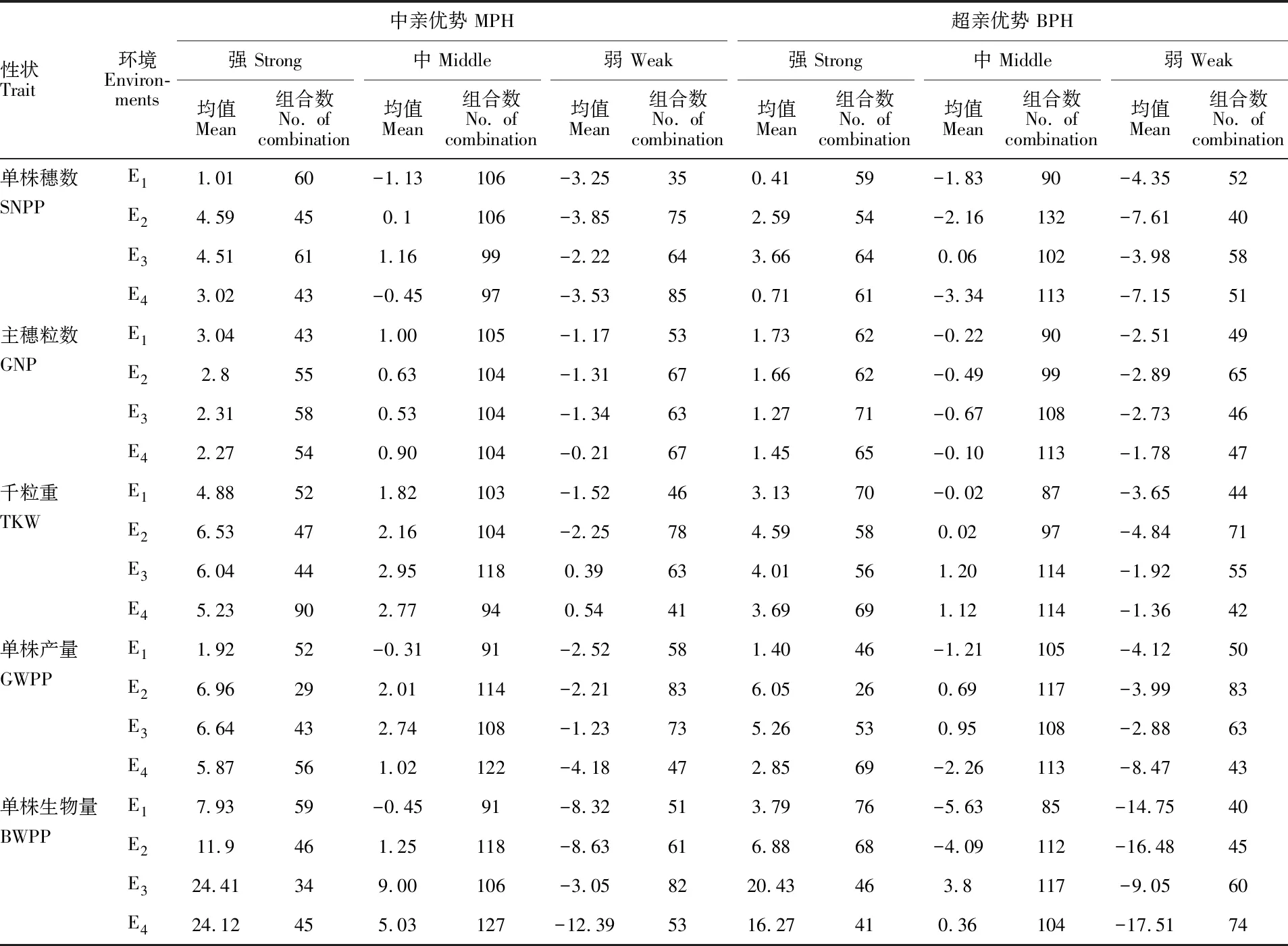
表7 大麥單株產量性狀中親優勢和超親優勢聚類分析Table 7 Cluster analysis of yield traits in MPH and BPH
2.5.3 大麥強優勢高穩定性組合的篩選
根據雜種優勢穩定性及強弱篩選強優勢高穩定性組合,結果(表8)發現,單數穗數、主穗粒數、千粒重、單株產量和單株生物量的強中親優勢高穩定性的組合分別為6、40、44、7和4個,強超親優勢高穩定性組合個數分別為2、15、26、4、和6個。就中親優勢而言,在226個雜交組合中,有9個強優勢且多環境穩定的大麥雜交組合,即100×101、093×095、106×107、008×010、013×014、041×042、057×059、096×097和012×013,其中100×101和057×059雜交組合為5個性狀均具有強超親優勢高穩定性的組合。
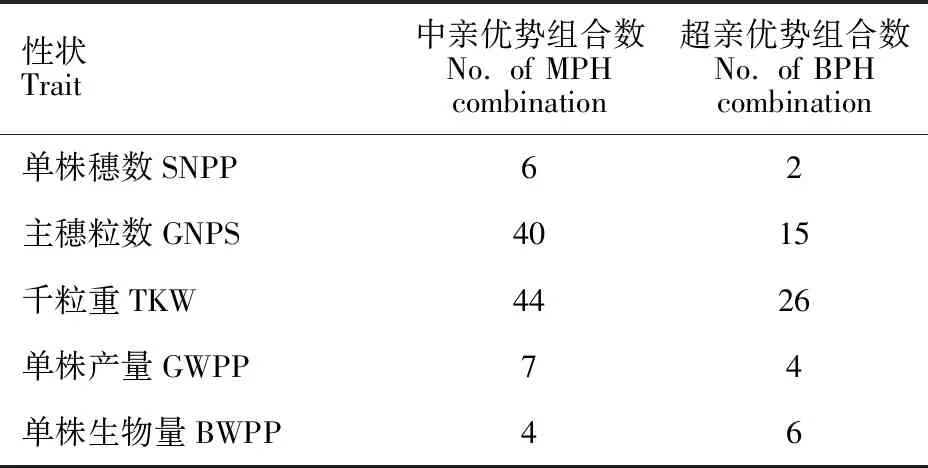
表8 大麥單株產量性狀強優勢高 穩定性組合的篩選Table 8 Screening of the superiority and high stability combination of yield traits in barley
3 討 論
3.1 大麥產量性狀雜種優勢表現的影響因素
雜種優勢利用是提高大麥產量的重要途徑之一,而雜種優勢的表現是由多基因控制的,同時受生長環境的影響。戴 維等[10]研究表明,不同環境下油菜含油量雜種優勢具有差異,但同一組合的雜種優勢表現具有較好一致性。邢朝柱等[11]研究認為,不同環境下雜交棉親本產量受環境影響較大,而雜交種產量相對穩定。顏秀娟等[13]研究表明,環境對大豆各性狀雜種優勢表現均有不同程度的影響,百粒重雜種優勢在不同環境間表現較為穩定。本研究結果表明,大麥中親優勢組合較為普遍,而超親優勢組合相對較少,大麥單株產量的雜種優勢因組合不同而異。大麥不同性狀的雜種優勢存在較大差異,千粒重雜種優勢最為顯著,單株生物量、單株產量、主穗粒數和單株穗數的雜種優勢相對較小。這與許如根等[5]和張新忠等[8]研究結果一致。方差分析結果表明,基因型和環境對大麥單株產量性狀均有顯著影響,說明大麥產量性狀雜種優勢受遺傳和環境因素共同作用,這與劉文江等[15]、劉麗華等[16]和趙延明等[17]在水稻、玉米等植物上的研究結果基本一致。
3.2 大麥產量性狀強優勢組合的評價
生產中理想的品種或作物雜交組合不但具有較強的雜種優勢而且具有較好的穩定性[18-20]。李 猛等[22]研究表明,不同玉米雜交種的產量在不同環境間穩定性的差異較大。蔣開封等[23]認為,雜交水稻的產量及其配合力和穩定性因雜交水稻組合不同而異,且穩產性的改良對豐產性的提高無顯著影響,培育既穩產又高產的雜交水稻是可行的。邢朝柱等[24]研究認為,轉基因抗蟲雜交棉產量性狀必須在生態環境中選擇。本研究表明,大麥單株產量單一性狀的雜種優勢表現不但因組合而異,同一組合的雜種優勢在不同環境間的變異也較大,穩定性好、雜交優勢強的組合較少。本研究根據雜種優勢穩定性及強弱篩選到5個性狀均具有強超親優勢高穩定性的組合100×101和 057×059。
猜你喜歡
礦山安全信息(2022年40期)2022-04-07 02:16:52
當代水產(2021年10期)2021-12-05 16:31:48
中老年保健(2021年12期)2021-08-24 03:30:40
中國傳媒大學學報(自然科學版)(2021年1期)2021-06-09 08:43:00
今日農業(2020年20期)2020-11-26 06:09:10
趣味(語文)(2020年3期)2020-07-27 01:42:46
中國生殖健康(2020年6期)2020-02-01 06:28:50
中國生殖健康(2019年11期)2019-01-07 01:28:02
聚氯乙烯(2018年9期)2018-02-18 01:11:34
作文與考試·初中版(2017年12期)2017-04-19 20:26:27
