干旱脅迫下過表達TaER小麥的轉(zhuǎn)錄組及光合特性分析
2019-08-22 09:54:20李慧娟鄭甲成胡銀崗
麥類作物學(xué)報 2019年8期
李慧娟,楊 陽,常 平,鄭甲成,陳 亮,胡銀崗
(西北農(nóng)林科技大學(xué)農(nóng)學(xué)院/旱區(qū)作物逆境生物學(xué)國家重點實驗室,陜西楊凌 712100)
小麥(TriticumaestivumL.)是僅次于水稻的世界第二大糧食作物,種植面積最大、分布最廣。在我國,由于小麥主栽區(qū)氣候特征的影響,小麥灌漿期降水量嚴重不足,最終導(dǎo)致大幅減產(chǎn),因此抗旱性成為小麥育種的重要指標,而提高蒸騰效率是抗旱性改良的關(guān)鍵。
ERECTA(ER)是調(diào)控植物蒸騰效率的1個編碼類受體蛋白激酶的基因,通過與大分子物質(zhì)的相互作用來調(diào)節(jié)植物的生長發(fā)育,并且在植物抵御病菌侵染、干旱、高溫等生物和非生物脅迫中也發(fā)揮重要作用[1]。Xing等[2]在擬南芥中過表達楊樹的ER基因,發(fā)現(xiàn)轉(zhuǎn)基因植株生物量、光合速率和長期水分利用效率都有顯著提高。Villagarcia等[1]發(fā)現(xiàn)在番茄中過表達缺失激酶催化結(jié)構(gòu)域的擬南芥ER基因,對生長造成負面效應(yīng)的同時降低了對水分虧缺的抗性。針對禾本科作物的研究表明,過表達ER基因的轉(zhuǎn)基因玉米、高粱和水稻也表現(xiàn)出相似的特性[3-4]。此外,Shen等[5]研究發(fā)現(xiàn),擬南芥ER基因?qū)χ参锏哪蜔嵝砸灿泻軓姷男?yīng)。盡管對擬南芥、水稻中ER基因的效應(yīng)有了比較深入詳細的研究,但是目前小麥TaER基因的功能研究尚處于起步階段,且其轉(zhuǎn)化相關(guān)研究還未見報道。
轉(zhuǎn)錄組測序技術(shù)是近年來興起的一種針對整個轉(zhuǎn)錄網(wǎng)絡(luò)進行定量分析的測序技術(shù),通過對不同樣品所有mRNA的高通量測序,既能夠分析不同樣品在特定生長發(fā)育時期所有基因的表達量,也可以分析相同組織在不同條件下的基因表達差異[6-7],已被廣泛應(yīng)用于植物應(yīng)對逆境響應(yīng)等研究。生物或非生物脅迫下,植物為了抵御逆境所造成的傷害,在基因的表達水平、分子代謝及生物代謝途徑方面都會發(fā)生極顯著的變化[8]。植物首先接收并識別由于環(huán)境變化而產(chǎn)生的信號,其次,在多種激酶及激素類物質(zhì)的作用下對信號進行傳導(dǎo),并通過調(diào)節(jié)相關(guān)基因的表達、轉(zhuǎn)錄后加工及翻譯水平來對逆境進行及時的響應(yīng),從而減少逆境對植物生長發(fā)育所帶來的傷害[9]。因此,轉(zhuǎn)錄水平上的改變是植物對環(huán)境響應(yīng)的第一步,通過對逆境下基因表達網(wǎng)絡(luò)和富集通路的深入分析有助于挖掘與逆境相關(guān)的重要基因及代謝途徑,從而進一步揭示植物響應(yīng)脅迫的調(diào)控機制。
小麥TaER對抗旱性的效應(yīng)及作用機制尚不明確,本研究以本實驗室前期構(gòu)建的過表達TaER基因的小麥T3代轉(zhuǎn)基因株系及其野生型為材料,利用轉(zhuǎn)錄組測序技術(shù),比較分析轉(zhuǎn)基因株系與野生型在干旱脅迫下的基因表達差異,通過功能富集,深入分析過表達TaER株系在干旱脅迫下所引起的基因表達調(diào)控網(wǎng)絡(luò),并測定其光合、蒸騰參數(shù),初步闡明TaER基因在干旱脅迫下的應(yīng)答機制,以期為小麥ER基因的功能研究奠定基礎(chǔ),促進小麥TaER基因應(yīng)用于小麥的抗旱性改良。
1 材料與方法
1.1 試驗材料與處理
以T3代轉(zhuǎn)基因小麥株系[以小麥栽培品種Cadenza為受體,將位于小麥7BS上的TaER基因(NCBI數(shù)據(jù)庫中的編號為JQ599261.2)的cDNA序列連接玉米組成型ubiqutin啟動子構(gòu)建表達載體,經(jīng)基因槍遺傳轉(zhuǎn)化獲得]通過目的基因檢測及表達水平分析獲得的高表達株系L1、L2及其野生型栽培品種Cadenza為材料。
試驗于2016-2017年間在西北農(nóng)林科技大學(xué)旱區(qū)農(nóng)業(yè)節(jié)水研究院抗旱棚內(nèi)(34°7′ N,108°4′ E)進行。根據(jù)完全隨機區(qū)組設(shè)計充分灌溉(well-watered,WW)和干旱脅迫(water-stressed,WS)兩種水分處理,每個處理設(shè)4次重復(fù)。
播種前充分灌溉,越冬、返青、拔節(jié)期,WW和WS處理依賴自然降水,期間的降水量為 184.2 mm。拔節(jié)后,抗旱棚平時開啟,雨天關(guān)閉,WS處理停止灌溉,WW處理分別在抽穗和灌漿初期各灌水20 mm。在灌漿中期分別取正常灌水與干旱脅迫下各株系的12個單株的旗葉樣品,其中4個單株旗葉樣品直接液氮速凍,-80 ℃超低溫冰箱保存用于DNA提取,其余8個單株的旗葉等量混樣速凍后用于轉(zhuǎn)錄組測序總RNA的提取。
1.2 TaER轉(zhuǎn)基因T3代陽性株系的鑒定
CTAB法提取各株系基因組DNA,在目的基因和載體區(qū)域分別設(shè)計正、反向引物對T3代轉(zhuǎn)基因株系的外源基因進行檢測,目的片段大小為354 bp,4個生物學(xué)重復(fù)。引物序列見表1。由上海生工生物工程有限公司合成。PCR反應(yīng)體系為10 μL:5.0 μL的2×taqPCR Star Mix,正、反向引物(10 μM)各0.5 μL,0.5 μL的模板DNA(100 ng·μL-1),ddH2O 3.5 μL。反應(yīng)程序為:94 ℃預(yù)變性2 min,94 ℃變性30 s,61 ℃退火30 s,72 ℃延伸20 s,35個循環(huán),最后72 ℃延伸 5 min。PCR產(chǎn)物使用1%的瓊脂糖凝膠電泳進行檢測,最終確定T3代轉(zhuǎn)基因陽性株系。
1.3 TaER轉(zhuǎn)基因株系的表達量分析
利用qRT-PCR檢測目的基因TaER的表達水平,使用Trizol法提取三葉期T3代轉(zhuǎn)基因陽性單株第一葉的總RNA并進行反轉(zhuǎn)錄。反應(yīng)總體積為20 μL,具體各組分參照SuperReal PreMix Color (SYBR Green) Kit熒光定量試劑盒說明書配置。反應(yīng)程序如下:95 ℃預(yù)變性 15 min,95 ℃變性10 s,60 ℃退火30 s,設(shè)置40個循環(huán),引物序列見表1。以TaActin為內(nèi)參基因,用2-△△Ct法計算相對表達量,每個樣品3次重復(fù)。

表1 PCR及qRT-PCR引物序列Table 1 Primer sequences for PCR and qRT-PCRW
1.4 轉(zhuǎn)錄組測序及表達量計算
用Trizol法提取總RNA,使用e-spect蛋白核酸分析儀對RNA質(zhì)量進行檢測,檢測合格的樣品送至北京諾禾致源科技股份有限公司進行后續(xù)建庫及測序。將測序獲得的序列去除接頭序列及低質(zhì)量reads之后,將clean data比對到水稻參考基因組上,使用HTseq進行表達量的計算。
1.5 差異表達基因的篩選及富集分析
用FPKM值統(tǒng)計基因的表達水平[10],用基于平均值的高斯分布和基于統(tǒng)計量的經(jīng)驗分布通過DEGseq[11]軟件進行差異基因的篩選,閾值為FDR≤0.01和│log2(fold change)│≥1,分別對充分灌水和干旱脅迫處理下的野生型與過表達株系的差異表達基因(DEGs)進行分析。
以q value <0.05為閾值,分別利用KOBAS3.0和GOseq[12]對差異基因進行GO(Gene Ontology)功能和KEGG(Kyoto Encyclopedia of Genes and Genomes)通路富集分析。
1.6 轉(zhuǎn)錄組數(shù)據(jù)的qRT-PCR驗證
為了驗證轉(zhuǎn)錄組數(shù)據(jù)的可靠性,以各處理葉片cDNA為樣品,選擇6個在不同水分處理下的差異表達的基因(表1),用qRT-PCR分析其表達量,3個重復(fù),具體反應(yīng)及程序參照1.3。
1.7 光合相關(guān)指標的測定
用LI-6400便攜式光合儀(Li-Cor,Lincoln/NE,USA)測定灌漿期各株系的光合、蒸騰參數(shù),同時參考Martin等的方法[13]測定并計算瞬時水分利用效率。用Excel進行數(shù)據(jù)的統(tǒng)計與整理,SPSS16.0進行方差分析與多重比較。
2 結(jié)果與分析
2.1 目的基因檢測及表達量分析
對T3代轉(zhuǎn)基因株系基因組DNA進行多次重復(fù)PCR檢測。結(jié)果顯示,轉(zhuǎn)基因陽性植株可以擴增出354 bp的特異片段,而非陽性植株和野生型則未擴增出目的條帶;15個株系中,有9個株系為陽性株系(圖1),陽性率為60%,表明部分T3代轉(zhuǎn)基因株系尚有分離。野生型標記為WT,T3代轉(zhuǎn)基因株系標記為L。
對獲得的陽性轉(zhuǎn)基因株系目的基因的表達量進行定量分析。結(jié)果表明,所有陽性轉(zhuǎn)基因株系中外源TaER基因的表達量都不同程度的上調(diào)(圖2),其中株系L1和L2上調(diào)幅度較高,因此以L1和L2株系為材料,進行轉(zhuǎn)錄組測序,進一步分析干旱脅迫下過表達TaER的轉(zhuǎn)基因株系與野生型在基因表達及調(diào)控網(wǎng)絡(luò)中的差異。
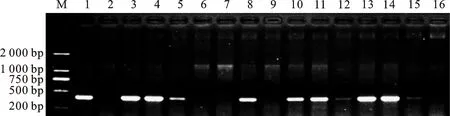
M:DNA Maker DL2000; 1~16分別為L1、WT、L2、L3、L4、L5、L6、L7、L8、L9、L10、L11、L12、L13、L14和L15。
M:DNA Maker DL2000; 1~16 represent L1,WT,L2,L3,L4,L5,L6,L7,L8,L9,L10,L11,L12,L13,L14 and L15,respectively.
圖1 轉(zhuǎn)基因植株TaER基因的PCR檢測結(jié)果
Fig.1 PCR analysis of transgenic plants forTaERgene
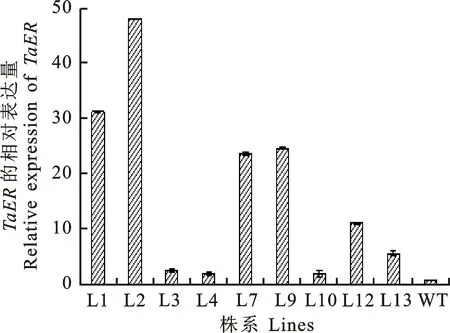
圖2 T3轉(zhuǎn)基因株系TaER基因的qRT-PCR檢測結(jié)果Fig.2 QRT-PCR analysis of TaER gene for T3 transgenic lines
2.2 測序數(shù)據(jù)的統(tǒng)計及比對情況評估
對2個水分處理下的野生型、L1和L2株系共6個混合樣品進行測序,經(jīng)過原始數(shù)據(jù)過濾、質(zhì)控之后,野生型(WT_WW、WT_WS)、轉(zhuǎn)基因株系L1(L1_WW、L1_WS)和轉(zhuǎn)基因株系L2(L2_WW、L2_WS)分別獲得76 217 622、61 641 432、61 915 306、74 464 998、79 375 080和61 391 468條clean reads,樣品平均GC含量60.44%,Q20比例為94.21%,Q30比例為86.62%,表明測序數(shù)據(jù)質(zhì)量良好,可進行下游分析。
將clean reads比對到水稻參考基因組上,結(jié)果(表2)顯示,6個樣品在基因組上都有比較高的比對率,平均比對率為87.76%,最低為87.08%,最高為89.73%。考慮到六倍體小麥遺傳背景的復(fù)雜性,超過87%的比對結(jié)果表明測序結(jié)果可靠,可以很好的滿足后期的分析需求。
2.3 差異表達基因分析
以FDR≤0.01和│log2(fold change)│≥1作為閾值,野生型為對照組,進行差異表達基因篩選,結(jié)果如表3所示,同時通過韋恩圖分析四組差異表達基因(圖3)。結(jié)果表明,充分灌溉下,有1 439個基因在2個轉(zhuǎn)基因株系和野生型之間均差異表達,其中470個上調(diào)表達,969個下調(diào)表達;干旱脅迫下,有607個基因表現(xiàn)出共同的差異表達,其中352個上調(diào)表達,255個下調(diào)表達。

表2 測序數(shù)據(jù)與參考基因組比對結(jié)果統(tǒng)計Table 2 Statistics of sequencing data mapped in reference genome

表3 過表達株系與野生型的差異表達基因數(shù)Table 3 Statistics of DEGs between over-expression lines and wild type
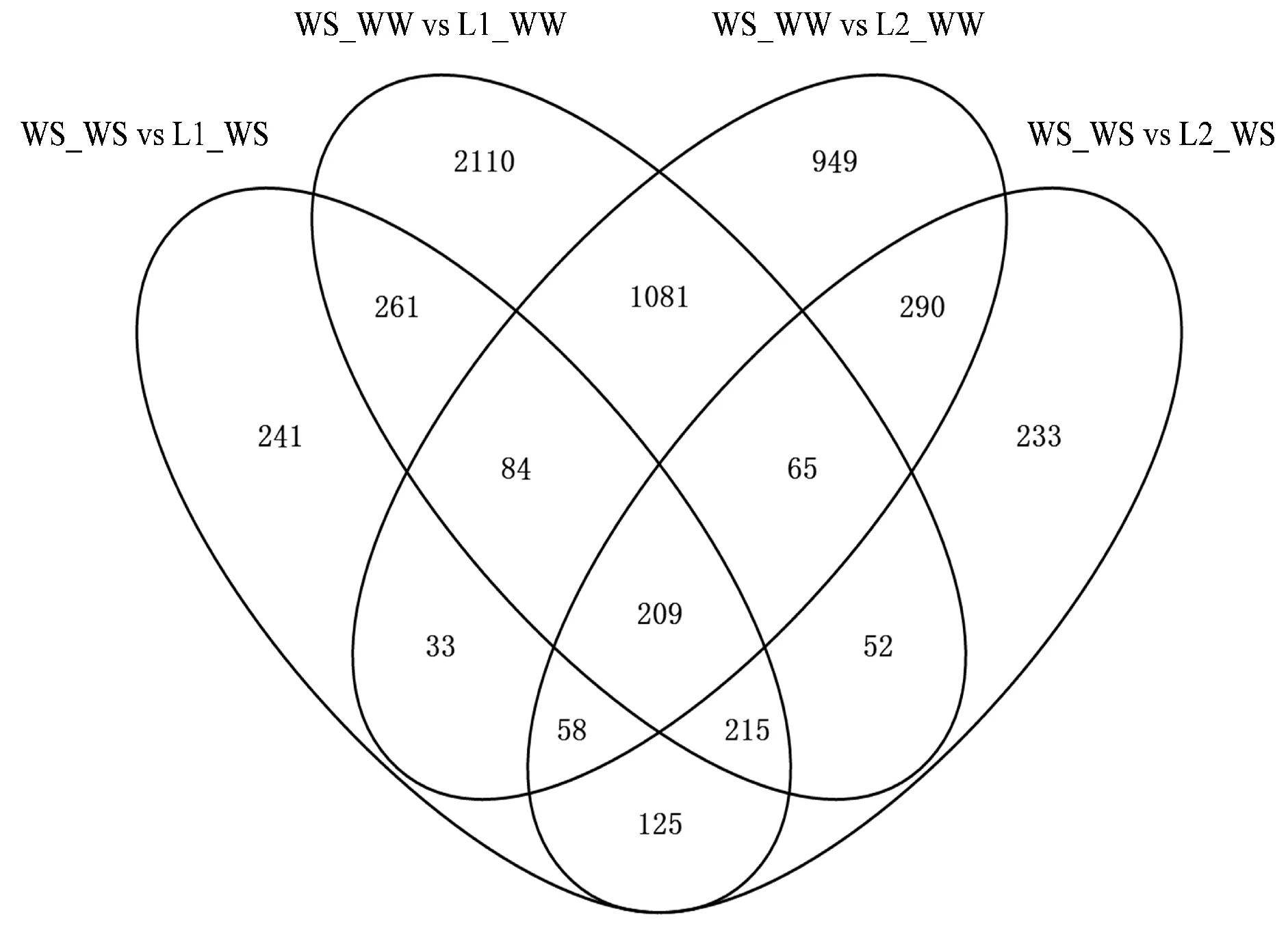
圖3 差異表達基因的韋恩圖Fig.3 Veen plots of DEGs
2.4 qRT-PCR驗證
為了驗證轉(zhuǎn)錄組表達數(shù)據(jù)的可靠性,選擇6個在干旱處理下野生型與轉(zhuǎn)基因株系顯著差異表達的基因,用qRT-PCR進行驗證。結(jié)果(圖4、圖5)表明,除了TRIAE_CS42_7DS_TGACv1_622753_AA2044770與轉(zhuǎn)錄組表達量數(shù)據(jù)有細微的差別外,其余基因都表現(xiàn)出與轉(zhuǎn)錄組數(shù)據(jù)一致的表達情況,表明轉(zhuǎn)錄組數(shù)據(jù)是客觀可靠的。
2.5 差異表達基因的GO和KEGG富集分析
2.5.1 差異表達基因的GO富集分析
對正常灌水處理下的1 439個DEGs進行GO功能分析,差異基因富集到分子功能、細胞組分和生物學(xué)過程3個類別的40個GO功能分類。其中,參與生物學(xué)過程的基因最多,其次為分子功能,較少的是細胞組分,分別占總數(shù)的58.65%、 28.05%和13.29%。富集差異基因數(shù)較多的20個GO功能(表4)中,細胞組分中類囊體和類囊體組的DEGs最多,均有28個,其次是光系統(tǒng)(24個)和光合膜(24個);分子功能中DEGs富集最多的是催化活性(502個)、水解酶活性(211個)和氧化還原酶活性(140個);生物學(xué)過程中代謝過程(548個)、光合作用(27個)的DEGs數(shù)量最多。
對干旱脅迫處理下的607個DEGs進行注釋,經(jīng)分析,DEGs富集到GO數(shù)據(jù)庫的生物學(xué)過程和分子功能2個類別,共有30個功能分類。其中,參與到生物學(xué)過程的DEGs占9.70%,參與到分子功能的占71.66%。富集差異基因數(shù)較多的20個GO功能(表5)中,生物學(xué)過程的DEGs較多富集在脂質(zhì)代謝(34個)和膜脂代謝(12個)過程;分子功能中主要是與催化活性(260個)和水解酶活性(139個)相關(guān)的基因。
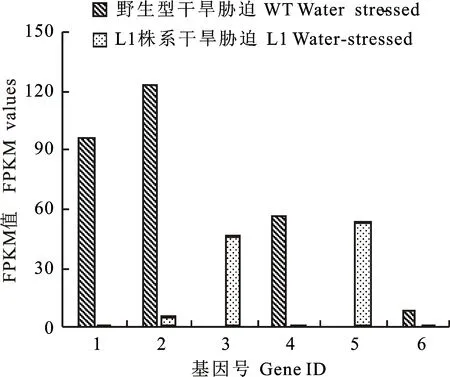
1~6:TRIAE_CS42_7DS_TGACv1_622753_AA2044770,TRIAE_CS42_7AS_TGACv1_569613_AA1820410,TRIAE_CS42_5BL_TGACv1_409850_AA1366530,TRIAE_CS42_4AL_TGACv1_288169_AA0939290,TRIAE_CS42_5BL_TGACv1_406947_AA1351690,and TRIAE_CS42_2BS_TGACv1_145841_AA0446640
圖4野生型和L1株系在干旱處理下差異基因的表達量FPKM值
Fig.4FPKM values of DEGs between wild lineand L1 line under drought stress
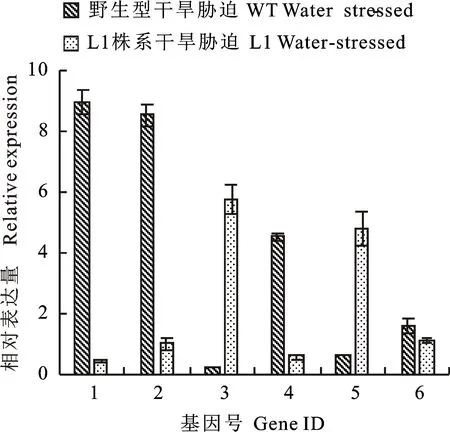
1~6:TRIAE_CS42_7DS_TGACv1_622753_AA2044770,TRIAE_CS42_7AS_TGACv1_569613_AA1820410,TRIAE_CS42_5BL_TGACv1_409850_AA1366530,TRIAE_CS42_4AL_TGACv1_288169_AA0939290,TRIAE_CS42_5BL_TGACv1_406947_AA1351690,and TRIAE_CS42_2BS_TGACv1_145841_AA0446640
圖5野生型和L1株系在干旱處理下差異基因的相對表達量
Fig.5Relative expression of DEGs between wildline and L1 line under drought stress

表4 正常灌水處理下野生型和過表達株系的差異表達基因GO注釋Table 4 GO annotation of DEGs under normal irrigation for wild type and over-expression lines
B、C、M分別為生物學(xué)過程、細胞組分、分子功能。表5同。
B:Biological process; C:Cellular component; M:Molecular function. The same in table 5.
2.5.2 差異表達基因的KEGG富集分析
結(jié)合KEGG數(shù)據(jù)庫,分析表明,正常灌水與干旱脅迫下,DEGs均顯著富集到8條途徑(表6)。干旱脅迫下,甘油磷脂、亞油酸、甘油脂和α-亞麻酸代謝途徑顯著富集,而光合作用-捕光蛋白及所涉及的3個氨基酸代謝途徑不再富集。
2.6 過表達株系和野生型的光合相關(guān)指標
在小麥灌漿期,測定正常灌水與干旱脅迫處理下過表達株系和野生型旗葉的凈光合速率與蒸騰速率(表7)。結(jié)果表明,正常灌水處理下,TaER過表達株系L1和L2的凈光合速率均顯著高于野生型,平均提高18.58%;蒸騰速率平均提高了21.24%;瞬時水分利用效率平均提高了19.94%。干旱脅迫處理后3個株系的凈光合速率與蒸騰速率均降低,但是過表達株系L1和L2的凈光合速率仍顯著高于野生型;蒸騰速率平均較野生型降低了8.83%;而瞬時水分利用效率平均為野生型的1.9倍。

表5 干旱脅迫處理下野生型與過表達株系差異表達基因的GO注釋Table 5 GO annotation of DEGs under drought stress for wild type and over-expression lines
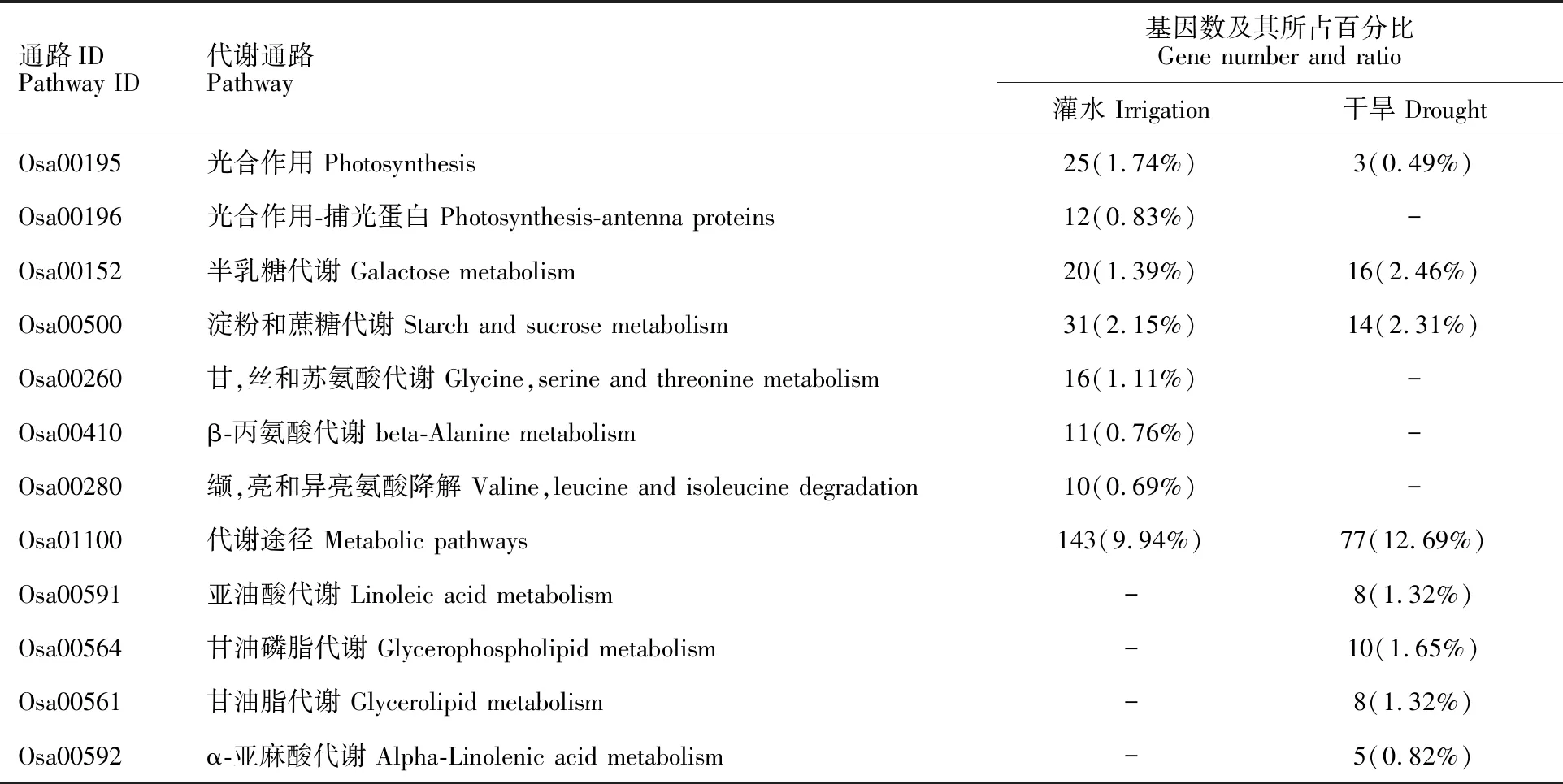
表6 野生型與過表達株系差異表達基因的KEGG富集分析Table 6 KEGG enrichment analysis of the DEGs between wild type and over-expression lines
-:無富集的差異表達基因。
-:Non-enriched differentially expressed genes.
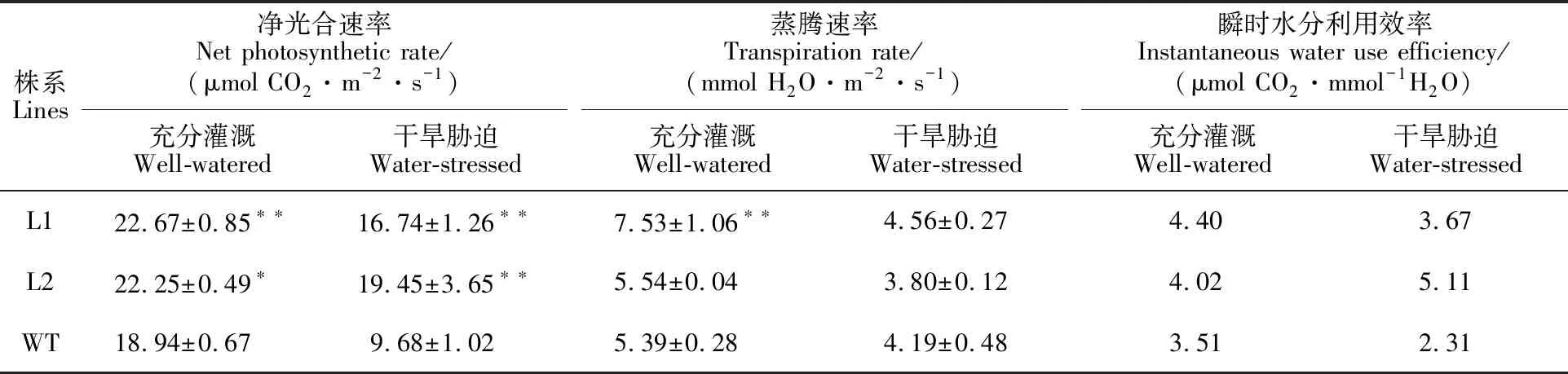
表7 灌漿期光合和蒸騰性狀分析Table 7 Analysis of photosynthesis and transpiration traits at grain-filling stage
*和**分別表示轉(zhuǎn)基因株系和野生型在0.05和0.01水平上差異顯著。
* and ** indicate significant differences between transgenic lines and wild type at 0.05 and 0.01 levels,respectively.
3 討 論
近年來,轉(zhuǎn)錄組水平的測序技術(shù)在很大程度上促進了生物基因功能方面的研究。如今,RNA-seq也廣泛應(yīng)用于植物應(yīng)對逆境的響應(yīng)。本研究通過對正常灌溉和干旱脅迫的2個T3代轉(zhuǎn)基因株系及其野生型旗葉進行RNA-seq測序,篩選出在2個轉(zhuǎn)基因株系與野生型之間正常灌水下的1 439個共同的差異表達基因,干旱脅迫下607個共同的差異表達基因。這表明某些基因參與小麥抗旱性的調(diào)控。而在正常灌溉條件下,L1和L2株系與野生型差異表達基因數(shù)目有較大的差異(L1為4 077個,而L2為2 769個),這可能是由于在基因轉(zhuǎn)化過程中插入位點、拷貝數(shù)的不同以及在基因重組互換過程中產(chǎn)生的差異而影響了其他基因表達的變化。目前對于小麥轉(zhuǎn)錄組學(xué)研究處于起步階段,基因功能注釋信息不夠完善,部分序列或基因無法獲得功能注釋信息,而水稻功能信息則比較完善,因此將水稻基因組作為參考進行比對。
正常灌水下DEGs的GO功能主要富集到光合作用、催化活性、水解酶活性和氧化還原酶活性、類囊體、類囊體組分,干旱脅迫下的DEGs富集到的主要是脂質(zhì)、膜脂代謝基因、催化活性和水解酶活性。表明光合作用、脂質(zhì)和膜脂代謝基因在干旱脅迫下顯著富集,暗示這些基因可能在TaER過表達株系在適應(yīng)干旱方面發(fā)揮著重要 作用。
本研究KEGG分析表明,干旱脅迫下,DEGs富集到半乳糖代謝、淀粉和蔗糖代謝、甘油(磷)脂代謝、亞油酸代謝、光合作用-捕光蛋白和α-亞麻酸代謝,與前人在禾本科早熟禾屬的結(jié)果基本一致[14]。干旱脅迫后的差異基因主要參與了脂類代謝和光合作用,且甘油酯途徑中的糖基轉(zhuǎn)移酶差異基因上調(diào)表達,擬南芥糖基轉(zhuǎn)移酶UGT75X被鹽脅迫、干旱脅迫誘導(dǎo)表達,通過糖基化脫落酸、油菜素內(nèi)酯、赤霉素、細胞分裂素和其他小分子物質(zhì)提高植物抗旱性[15]。表明這些基因可能通過植物激素調(diào)節(jié)對干旱的適應(yīng)。亞油酸和α-亞麻酸代謝中的脂氧合酶(LOX)基因下調(diào)表達。其參與生成脫落酸、茉莉酸等抗性信號分子響應(yīng)干旱脅迫,也可以催化亞麻酸氫過氧化而抑制光合電子的傳遞[16]。因此,這些基因可能通過植物激素、光合作用正調(diào)控干旱脅迫。光合作用-捕光蛋白途徑中Lhca1、Lhcb(1、3、4和6)差異表達基因在充分灌溉下均下調(diào)表達,而干旱脅迫后該途徑則無差異表達基因的富集,這些基因參與光吸收和光信號轉(zhuǎn)導(dǎo),是光系統(tǒng)的組成部分[17]。在擬南芥光合作用調(diào)控中,ERECTA通過影響羧化酶最大羧化速率和電子傳遞能力來影響其光合能力[18],推測這些基因可能通過調(diào)控光電子傳遞途徑,從而適應(yīng)水分逆境。
植株的凈光合速率可以作為評價其抗旱性強弱的生理指標之一,抗旱性強的品種,能夠維持較高水平的光合特性[19]。分析田間光合、蒸騰參數(shù),結(jié)果表明TaER過表達提高了轉(zhuǎn)基因小麥株系的光合作用,這與擬南芥中ERECTA過表達提高凈光合速率的結(jié)果一致[18],且在干旱脅迫下維持較高的凈光合速率與瞬時水分利用效率,降低蒸騰速率。因此,干旱脅迫下,TaER過表達株系可能通過調(diào)控光合作用和脂類代謝等途徑提高凈光合速率,為TaER調(diào)控蒸騰效率和響應(yīng)干旱脅迫提供了理論基礎(chǔ)。
猜你喜歡
音樂探索(2022年2期)2022-05-30 21:01:37
民用飛機設(shè)計與研究(2020年4期)2021-01-21 09:15:02
小天使·一年級語數(shù)英綜合(2019年8期)2019-08-27 02:23:00
中國特種設(shè)備安全(2018年11期)2019-01-08 02:08:32
電子制作(2018年18期)2018-11-14 01:48:24
小學(xué)科學(xué)(學(xué)生版)(2018年7期)2018-08-13 09:33:04
山東工業(yè)技術(shù)(2016年15期)2016-12-01 05:31:22
鄭州大學(xué)學(xué)報(醫(yī)學(xué)版)(2015年2期)2015-02-27 14:50:46
中國中醫(yī)藥現(xiàn)代遠程教育(2014年11期)2014-08-08 13:23:44
山東女子學(xué)院學(xué)報(2014年6期)2014-03-01 02:24:55
