氮限制和海洋酸化對顆石藻Emiliania huxleyi NIWA1108生理指標的交互影響
2019-08-22 01:55:06馮媛媛李文學李敬鴻倪紅東石文婷
天津科技大學學報 2019年4期
廖 晏,馮媛媛,劉 瑤,李文學,李敬鴻,倪紅東,石文婷
(天津市海洋資源與化學重點實驗室,天津科技大學海洋與環境學院,天津 300457)
工業革命以來,人類活動的加劇導致大氣中二氧化碳(CO2)排放量急劇上升,到21 世紀末,CO2濃度將超過800μatm[1].海洋吸收了人為排放CO2的1/3[2],大量CO2被海洋持續吸收,導致海水pH 降低,被稱為海洋酸化(ocean acidification)[3].研究[4-5]表明,海洋酸化可影響海洋浮游植物的生長,尤其是對于具有鈣化作用的藻類[6-8].此外,CO2是溫室氣體的重要組成部分,對全球氣候起到重要的調節作用.隨著CO2濃度不斷升高,溫室效應增強使得溫度不斷升高,導致冰川融化和降雨模式發生改變,從而增強海水分層、減低海洋混合層深度[9]、削弱從深海向表層海水的營養鹽補充、加劇大洋表層水體的營養鹽限制[10].
海洋顆石藻是一類生產碳酸鈣(CaCO3)顆石粒作為外殼的單細胞浮游植物[11],它們能夠通過光合作用和鈣化作用進行有機碳和無機碳(CaCO3)的生產[12],貢獻了全球海洋CaCO3生產量的50%,在海洋碳循環中起重要作用[10,13-15].除極地及熱帶海域外,顆石藻(Emiliania huxleyi)在全球海洋中都有廣泛分布,并在很多海域中出現大規模可通過衛星遙感圖像觀測到的季節性水華[16-18],成為海洋碳循環研究中的模式生物[18].近年研究[19]表明,Emiliania huxleyi 的鈣化作用對海水碳酸鹽化學體系的變化響應尤為敏感,并存在明顯的株間特性.除海洋酸化外,其生長、鈣化作用及光合作用等生理學過程還受到其他環境因子的影響,如光照、溫度、營養鹽濃度變化等[19-23].因此,研究海洋酸化和其他環境因子的交互作用對顆石藻生理學指標的影響,對進一步預測未來多因子共同變化的海洋環境中Emiliania huxleyi 的生理學響應尤為重要.氮是維持浮游植物生長的一種重要的生源要素,也是在全球大洋水體中限制浮游植物初級生產的重要營養鹽,然而針對海洋酸化和氮限制的交互作用對Emiliania huxleyi 的生理學指標影響的相關研究,尤其是在穩態生長下該藻的生理學響應卻鮮有報道[20,24].本研究擬采用恒化培養裝置對顆石藻Emiliania huxleyi NIWA1108 進行實驗室內受控連續培養實驗,研究海洋酸化和氮限制對該藻生理學指標在其穩態生長條件下的交互影響效應.
1 材料與方法
1.1 實驗設置
本實驗所用顆石藻是顆石藻鈣化株Emiliania huxleyi NIWA1108,于2009 年分離自新西蘭以東Chatham Rise 海域(41°35.8′S,175°41.5′E),在實驗室中用f/20 海水培養基(10 倍稀釋后的f/2 配方)進行保種培養[25],培養條件為 15 ℃、光照強度 80~110μmol/(m2·s)、光/暗周期為12 h/12 h.配制培養基所使用的海水取自于南海寡營養海域SEATS 站表層,并在使用前通過0.2μm 孔徑的囊式濾器進行過濾處理,以達到無菌效果.
氮限制培養實驗采用自制恒化培養器進行,共設置4 個處理組:(1)對照:p(CO2)400μatm,氮充足(n(N)/n(P)=24);(2)酸化:p(CO2)800μatm,氮充足;(3)氮限制(n(N)/n(P)=2.4):p(CO2)400μatm,低氮;(4)酸化+氮限制:p(CO2)800μatm,低氮.培養基采用過濾后SEATS 站表層海水配制,磷酸鹽、微量元素及維生素添加至f/20 配方水平.氮充足條件下的培養基硝酸鹽濃度添加至 f/20 水平(88μmol/L);低氮條件下硝酸鹽濃度添加至8.8μmol/L(n(N)/n(P)=2.4).初始接種豐度為1.0×104mL-1,采用恒化培養器在培養箱內進行連續培養,至接種后第3 天打開蠕動泵,開始連續培養實驗.培養基采用蠕動泵連續泵入培養瓶(培養瓶體積為3.6 L,為聚碳酸酯材質)中,每個培養瓶瓶頸處與流出口相連,以保證培養體系體積恒定.蠕動泵的稀釋速率對于氮充足和氮限制處理組分別設置為0.5 d-1和0.2 d-1.實驗過程中使用具有特氟龍涂層的攪拌器連續低速攪拌,使得藻液分布均勻.培養實驗在恒溫光照培養箱中進行,溫度及光照條件與保種條件保持一致.
1.2 海水碳酸鹽體系調節
采用曝氣法調節海水培養基的碳酸鹽體系.其中空氣對照組(p(CO2)400μatm)向其中一桶海水中曝空氣(空氣中平均p(CO2)400μatm,從實驗開始曝氣直到實驗結束),另一桶海水中通入空氣和CO2混合氣體(p(CO2)800μatm,從實驗開始通氣直到實驗結束),氣體在通入水體前均經0.22μm 的濾膜過濾.培養開始前,海水培養基曝氣時間大于24 h,且在培養期間,儲存海水培養基的儲水桶和培養瓶一直保持持續曝氣.各培養瓶中在細胞生長達到穩態后的碳酸鹽體系各參數見表1,采用CO2SYS 1.05 版軟件進行計算[26].
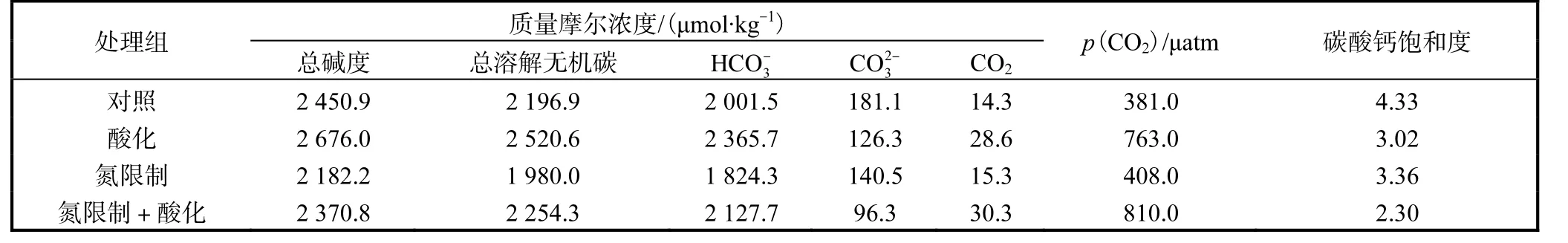
表1 采樣時恒化器中碳酸鹽化學體系參數Tab.1 Carbonate chemistry parameters in the chemostat vessels at the time of sampling
1.3 藻種培養
將處于指數生長期的 Emiliania huxleyi NIWA1108 接種于3.5 L 的培養液中,初始細胞豐度為1.0×104mL-1,置于光照培養箱中連續培養15 d.每隔24 h 取5 mL 藻液,分成3 份:一份用Trilogy 熒光儀測定活體熒光值;一份用pH 計測定pH;最后一份固定后用于細胞計數.培養至其生長進入穩態超過5 d(每個處理組中藻細胞豐度保持相對穩定,變化小于10%)后,進行最終采樣,采樣參數為活體熒光、pH、細胞計數、葉綠素a(Chl-a)、沉降速率及細胞元素組成.細胞元素組成包括顆粒有機磷(POP)、顆粒有機碳(POC)、顆粒無機碳(PIC)及顆粒有機氮(PON)含量.
1.4 樣品分析
1.4.1 藻細胞計數及Chl-a 質量濃度的測定
取 1 mL 藻液并加入 6 μL 堿性魯格氏溶液(Lugol’s)[27],并在4 ℃于暗處保存,最后用微藻計數框在顯微鏡下觀測計數.取樣至最后觀測時間間隔不超過一周.
量取 30 mL 藻液,用隔膜真空泵過濾到Whatman GF/F 玻璃纖維膜上(直徑25 mm),并保存于-20 ℃的冰箱中.測定時,在暗處將膜置于20 mL棕色玻璃瓶中,添加5 mL 體積分數為90%的丙酮,于-20 ℃暗處理24 h 后,用 Turner 型熒光儀測定其熒光值,根據式(1)[28]計算Chl-a 質量濃度(μg/L).

1.4.2 細胞元素組成的分析
POP 含量用鉬酸鹽測定法[29]進行測定.取30 mL 藻液過濾到經馬弗爐灼燒(450 ℃,4 h)過的GF/F 膜上,用2 mL 0.17 mol/L 的Na2SO4溶液潤洗,轉移到經馬弗爐灼燒(450 ℃,4 h)的硼酸鹽閃爍瓶中,用2 mL 0.017 mol/L MgSO4溶液浸沒附有藻液的GF/F 膜,然后將其置于60 ℃烘箱中,直至烘干.分析前,將裝有GF/F 膜的閃爍瓶置于馬弗爐中450 ℃灼燒4 h,冷卻后向其中加入5 mL 0.2 mol/L 鹽酸,旋緊瓶蓋置于90 ℃烘箱中烘30 min.冷卻后,加入染色劑[29],靜置10~20 min,用紫外-可見分光光度計測定吸光度.過濾3 份等體積的培養基,作為空白樣.
POC、PIC 和PON 含量用CHN 元素分析儀進行測定.取2 份100 mL 藻液分別過濾到事先經馬弗爐灼燒(450 ℃,4 h)過的GF/F 膜上,一份不進行酸化處理,直接在60 ℃下烘干,用于測定總顆粒碳(TPC)含量;另一份用濃鹽酸熏蒸3 h 后置于烘箱中60 ℃烘干,用來測定POC 和PON 含量.TPC 和POC 的差值即為PIC 含量.
1.4.3 沉降速率的測定
同源性采樣法[30]用于測定浮游植物沉降速率.將沉降柱豎直固定在支架上,堵住3 個出水口,把藻液混勻,倒入沉降柱中,使藻液充滿沉降柱,蓋好沉降柱使其密封(避免氣泡),在相同溫度下避光靜置2~4 h.從上到下分層取樣,記錄各層藻液體積,并分別過濾至GF/F 膜上,保存于-20 ℃的冰箱中,用于測定其葉綠素生物量.根據Bienfang[30]的沉降公式計算沉降速率.
1.4.4 掃描電鏡圖像觀察
取1~5 mL 藻液過濾至0.6μm 聚碳酸酯濾膜上,過濾時控制壓力小于0.02 MPa,再將濾膜輕輕平鋪于培養皿中,自然風干.最后,用JSM-IT300 型掃描電子顯微鏡進行形態觀察并拍照.
1.5 數據分析
環境因子的交互效應按照Folt 等[31]方法計算,單一環境因子(酸化(A)或氮限制(N)或兩環境因子共同變化(A+N)下對某生理生化參數的表觀影響(observed effect,OE)按照該處理組與對照組的變化百分比計算而得(正值為正向升高效應,負值為負向降低效應),二者的交互效應(multiplicative effect,ME)按照公式[31]:MEA+N=(1+OEA)×(1+OEN)-1計算.當|OEA+N|>|MEA+N|時,兩種環境因子為協同性交互效應;當|OEA+N|<|MEA+N|時,兩種環境因子為對抗性交互效應.由于本實驗采用恒化連續培養方式,每個處理組到最終采樣時已在各培養條件下保持若干代的穩定生長狀態,沒有平行樣帶來的誤差,因此可以得到相應降低[32].
2 結 果
2.1 細胞Chl-a 含量
不同條件下Emiliania huxleyi NIWA1108 的細胞Chl-a 含量如圖1 所示.當硝酸鹽濃度一致,不同CO2分壓條件下Emiliania huxleyi NIWA1108 胞內Chl-a 含量變化較小.但是在氮限制的條件下,p(CO2)400μatm 處理組中胞內Chl-a 含量與氮充足條件下相比降低了58.9%.在高二氧化碳分壓和氮限制的共同作用下,胞內的Chl-a 含量進一步降低,比p(CO2)400μatm、氮充足的對照組降低了66.2%.
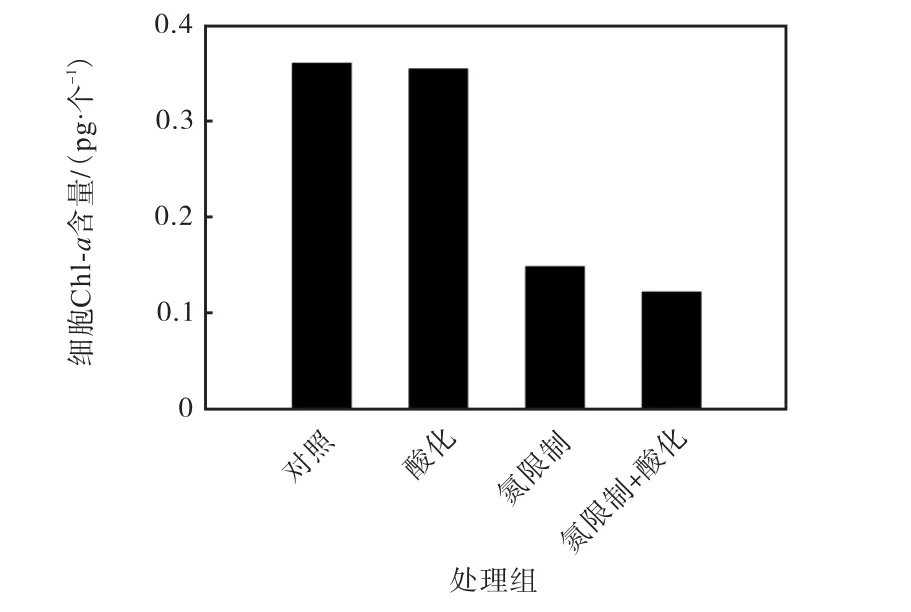
圖1 不同條件下Emiliania huxleyi NIWA1108 的細胞Chl-a 含量Fig.1 Contents of cellular chlorophyll a in Emiliania huxleyi NIWA1108 under different conditions
2.2 細胞元素含量
Emiliania huxleyi NIWA1108 的細胞POP 含量受到CO2分壓升高和氮限制的影響(表2).在氮充足的條件下,僅升高CO2分壓,細胞POP 含量比對照組降低23.1%;在氮限制條件下,細胞POP 含量比對照組降低了48.8%;高CO2濃度和氮限制的共同作用下,細胞POP 含量進一步降低,降低了65.4%.
與細胞POP 含量相似,細胞PON 含量隨CO2分壓升高有下降的趨勢(表2),而氮限制則進一步降低了該含量.在酸化和氮限制的共同作用下細胞內PON 含量下降程度最高(與對照組相比降低了54.0%).
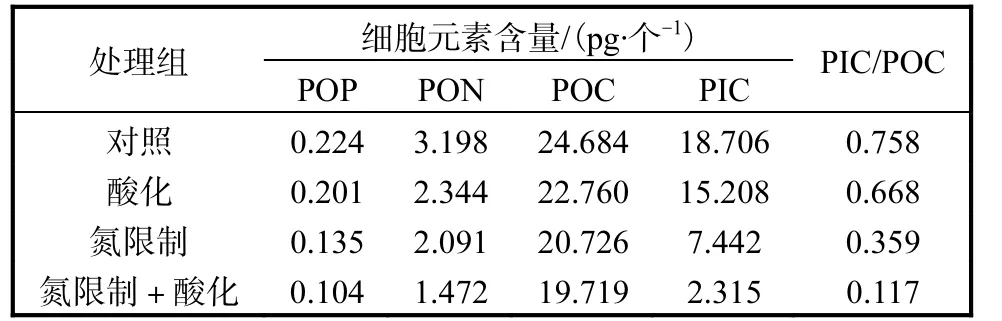
表2 不同條件下Emiliania huxleyi NIWA1108 的細胞元素含量Tab.2 Cellular elemental contents of Emiliania huxleyi NIWA1108 under different conditions
二氧化碳分壓的升高及氮限制均使細胞POC 含量略有下降(表2),與對照組相比,分別降低7.8%和16.0%,而酸化+氮限制組則進一步降低,較對照組低20%.而PIC 含量受海水酸化以及氮限制的影響比較大(表2),酸化組中Emiliania huxleyi NIWA1108的細胞PIC 含量與對照組相比降低了18.7%;氮限制使得PIC 含量大大降低(與對照組相比降低60.2%);氮限制和海水酸化的共同作用下,進一步降低了PIC含量(與對照組相比降低87.6%).
與對照組相比,僅升高CO2分壓的酸化組中,細胞PIC 與POC 比值(PIC/POC)有所降低(表2),降低了11.8%,氮限制下PIC/POC 下降得更為明顯,氮限制處理組中PIC/POC 較對照組降低了52.6%;在酸化+氮限制處理組,細胞PIC/POC 最低,比對照組降低84.5%,比酸化組降低82.4%.
2.3 沉降速率
高CO2分壓條件下(酸化組)Emiliania huxleyi NIWA1108 的沉降速率較對照組略有降低,而氮限制更加明顯地降低了其沉降速率,氮限制及酸化+氮限制處理組分別較對照組下降47.9%和48.4%(圖2).
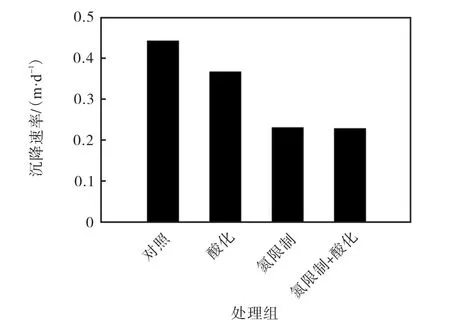
圖2 不同條件下Emiliania huxleyi NIWA1108 的沉降速率Fig.2 Sinking rates of Emiliania huxleyi NIWA1108 under different conditions
2.4 細胞形態
不同條件下Emiliania huxleyi NIWA1108 的掃描電鏡照片如圖3 所示.

圖3 在不同條件下Emiliania huxleyi NIWA1108 的掃描電鏡照片Fig.3 SEM images of Emiliania huxleyi NIWA1108 under different conditions
與對照組相比,Emiliania hux-leyi NIWA1108 的顆石球體積在高二氧化碳分壓和氮限制條件下均有所降低,在氮限制條件下細胞相對更小且顆石粒脫落更多;在酸化+氮限制處理組,其顆石球體積最小、顆石粒直徑也較小且存在大量顆石粒脫落.
2.5 海洋酸化和氮限制的交互作用
酸化和氮限制對顆石藻 Emiliania huxleyi NIWA1108 的各項生理及生物地球化學指標的交互作用見表 3.數據分析表明,酸化和氮限制對Emiliania huxleyi NIWA1108 細胞Chl-a 含量、PIC 含量、PIC/POC、細胞PON 含量及POP 含量產生協同性交互效應,而酸化和氮限制對細胞POC 含量和沉降速率產生對抗性交互效應.
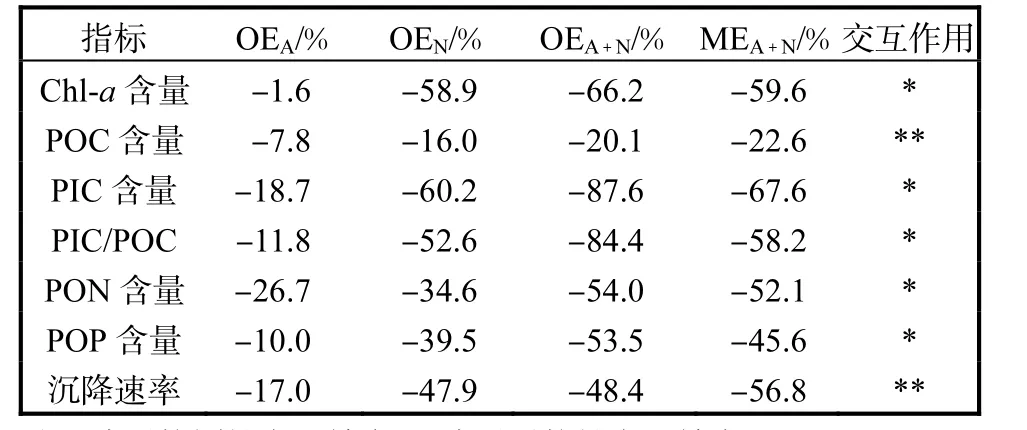
表3 酸化和氮限制對顆石藻Emiliania huxleyi NIWA1108的各項生理及生物地球化學指標的交互作用Tab.3 Interactive effects of ocean acidification and nitrogen limitation on different physiological and biogeochemical parameters of Emiliania huxleyi NIWA1108
3 討 論
本研究采用了恒化連續培養(chemostat continuous incubation)的方式[33],研究該藻在指數生長階段的穩態條件下氮限制、酸化以及二者的交互作用對其生理指標的影響,更好地模擬自然低硝酸鹽水體中顆石藻 Emiliania huxleyi NIWA1108 的自然生長狀態.這與報道的一次性培養實驗(batch culture)有所不同[34],一次性培養實驗達到的氮限制條件往往是在營養鹽消耗殆盡后的靜止平臺生長期,藻細胞瀕于衰亡,更與藻類發生大規模水華后處于衰退期的狀態相近.
氮限制可明顯抑制Emiliania huxleyi NIWA1108的生長、光合作用、鈣化作用,并降低其沉降速率,表明硝酸鹽濃度是控制該藻生理生化過程的重要環境因子.同樣,Feng 等[23]也發現硝酸鹽濃度在Emiliania huxleyi NIWA1108 生長、光合及鈣化作用中起到的重要作用,且在5 種環境因子中最為重要.氮是合成核酸和蛋白質的必需元素,氮限制可能會減少一些作為光合作用和鈣化作用的重要轉運體的蛋白質的生產[35].在本研究中,氮限制使Emiliania huxleyi NIWA1108 的細胞大小減小,因而細胞POC和PIC 含量均有所降低,然而氮限制下細胞PIC 降低的幅度更大,從而導致了細胞PIC/POC 比值在氮限制下明顯下降.這與Müller 等[36]和Perrin 等[37]研究結果有所不同,其研究對象為處于靜止生長期的Emiliania huxleyi NIWA1108 的細胞而無論在低光照或者高光照條件下,氮限制增加了細胞PIC/POC 比值,主要原因是處于靜止期的藻細胞在細胞生長的G1 期,鈣化作用繼續而細胞分裂暫停,導致細胞PIC含量相對升高,本研究對象為指數生長期藻細胞,在極低硝酸鹽濃度下,其鈣化作用較細胞分裂更大程度受到抑制.氮限制導致的細胞體積減小和PIC 含量降低也大大降低了其相對于海水的比密度,從而導致沉降速率降低,這將會進一步降低該藻向深海的碳沉降通量.同樣的,本研究中,隨著細胞變小,細胞Chla 含量也減少,細胞PON 含量降低[23,38].同時由于功能性蛋白合成受到氮限制影響,降低對營養鹽的吸收與有機磷合成,導致細胞POP 含量也降低.
本研究進一步表明了酸化和氮限制對顆石藻的生理和生物地球化學指標的潛在交互影響.酸化會導致顆石藻細胞花費更多的能量維持細胞內pH 平衡[39],因此隨著 CO2分壓升高,Emiliania huxleyi NIWA1108 的鈣化作用通常會隨之減弱[7,35].本研究中,當p(CO2)由400μatm 升高至800μatm 時,細胞PIC 含量和PIC/POC 比值均有所降低;而這種趨勢在氮限制下尤為明顯,換句話說,酸化和氮限制的交互作用對Emiliania huxleyi NIWA1108 的鈣化作用產生了負向協同效應.此外,酸化和氮限制也對Emiliania huxleyi NIWA1108 的細胞碳氮比和碳磷比產生了正向協同效應.考慮到在未來全球變化下的海洋環境中,除了海洋酸化的趨勢外[40],寡營養海域尤其是氮限制海域也會逐漸擴張[41],因此,為了更加準確地預測全球變化下顆石藻的生理學響應及該響應帶來的對海洋生物地球化學尤其是碳循環的影響,酸化和氮限制的這種協同影響不容忽視.
4 結 論
通過對顆石藻Emiliania huxleyi NIWA1108 進行恒化連續培養,揭示了在穩態生長條件下,氮限制對該藻的生理學指標影響較大,減小了顆石球的體積,降低了細胞各元素含量;而酸化則進一步降低了細胞元素含量,尤其是細胞無機碳含量以及PIC/POC 比值,其沉降速率也隨之降低.酸化和氮限制對顆石藻生理及生物地球化學指標尤其是鈣化作用存在著潛在協同效應,這為進一步預測全球變化下顆石藻這一重要功能群的響應以及氣候變化相關海洋生物地球化學模型的建立提供了理論支撐.
